ПРОБЛЕМЫ И ПЕРСПЕКТИВЫ КУЛЬТИВИРОВАНИЯ ДВУСТВОРЧАТЫХ МОЛЛЮСКОВ В БАРЕНЦЕВОМ МОРЕ
Увеличение антропогенного пресса на водоемы,
перелов многих промысловых гидробионтов имеют своим
следствием сокращение естественных ресурсов рыб
и водных беспозвоночных. В полной мере это относится
к арктическим морям, где в последние десятилетия XX в.
в неудовлетворительном состоянии оказались запасы
многих видов промысловых рыб: сельди, мойвы,
атлантического лосося, большинства сиговых и др.
В связи с кризисными явлениями в популяциях главных
промысловых объектов усилилось давление промысловиков на "дополнительные" объекты
промысла: пикшу, морского окуня, камбаловых рыб. В традиционных местах промысла резко
сократилась площадь зарослей бурых водорослей. Кризисные явления охватили поселения
моллюсков. В Белом море отмечаются случаи гибели мидиевых банок. Практически
нерентабельным стал промысел гребешков [1].
Резкое сокращение биоресурсов обусловливает необходимость развития аквакультуры,
под которой понимается получение биологической продукции путем культивирования
Работа выполнена за счет Программы Президиума РАН I.32П "Поисковые фундаментальные
научные исследования в интересах развития Арктической зоны Российской Федерации" в рамках темы
"Биоресурсы арктических морей России: современное состояние, влияние природных изменений
и антропогенных воздействий, научные основы и перспективы использования.
(выращивания) водных организмов ― гидробионтов [1]. В настоящее время марикультура
двустворчатых моллюсков в Баренцевом море делает первые шаги, хотя, например для Белого
моря, детально разработаны методики культивирования мидий [1].
В представленном обзоре даются краткие сведения по биологии потенциальных объектов
аквакультуры, относящихся к двустворчатым моллюскам (мидия съедобная и исландский
гребешок), и обобщаются имеющиеся наработки российских ученых, которые могут быть
полезны при организации акваферм, направленных на выращивание ценных промысловых видов
гидробионтов.
Мидия Mytilus edulis L. (рис. 1) является одним из наиболее распространенных донных
беспозвоночных прибрежья Баренцева и Белого морей. На каменистых и скальных грунтах
литоральной зоны, а иногда и в сублиторали, эти моллюски становятся доминирующим
или характерным видом многих биоценозов. Их биомасса составляет порой несколько
килограммов на 1 м2. Эврибионтность, доступность сбора, хорошая скорость роста, простота
выращивания и вкусовые качества мяса сделали мидий с давних пор объектом промысла
и культивирования, в том числе и на Севере. Опыт экспериментального выращивания
на Восточном Мурмане показал их несомненную перспективность для полярной аквакультуры [2].

Рис. 1. Мидия Mytilus edulis
Мидии обладают очень высокой толерантностью к колебаниям факторов среды. Взрослые
особи способны в течение нескольких часов осушения переносить температуры от -15 до +30 °С
[3] и соленость от 2.4 до 50 ‰ [2], но в воде с содержанием солей менее 5 ‰, не имея
предварительной адаптации, довольно быстро погибают [3].
Тело мидии заключено в выпуклую раковину, состоящую из двух практически равных
створок треугольно-овальной формы. Раковина темно-оливковая, темно-коричневая, чаще
черная. Нередко рисунок раковины дополняют чередующиеся темные и светлые радиальные
полосы, особенно хорошо заметные на просвет створок светлых тонов.
Проблемы и перспективы культивирования двустворчатых молюсков в Баренцевом море
ВЕСТНИК Кольского научного центра РАН 3/2016(26) 59
Форма и цвет раковины варьируют в зависимости от мест обитания. Передний конец
раковины совпадает с макушкой и немного заострен. Это составляет отличительную особенность
мидий. Брюшной край створок прямой или слабоизогнутый, спинной край сильно изогнут.
Задний конец раковины равномерно закруглен. Замочный край без зубов, но часто под макушкой
имеется несколько мелких зубчиков.
Тело мидии состоит из туловища и небольшой ноги. Головной отдел полностью
редуцирован. На переднем конце туловища лежит рот, на заднем ― порошица. Туловище
покрыто большими боковыми складками ― мантией. Они отходят от передней части туловища,
охватывают все тело и срастаются на спине. Пространство между складками и туловищем носит
название мантийной полости, где расположены жабры и органы пищеварительной, кровеносной,
выделительной, половой и нервной систем мидии. Сзади находится небольшой выводной сифон,
через который вода выходит из мантийной полости. Сквозь мантию проходят мускулы-
аддукторы раковины и нога. Биссус, при помощи которого моллюск прикрепляется
к субстрату, представляет собой тонкие прочные нити рогоподобного вещества. Биссусная
железа, выделяющая эти нити, находится в основании ноги. Жабры расположены в задней
половине тела и представлены жаберной осью с жаберными нитями. Жабры не только
обеспечивают ток воды, но и являются фильтрующим органом. Приносимые частицы взвеси
сортируются здесь по размеру и качеству. Непригодные к поглощению частицы направляются
к выводному сифону и удаляются из организма в виде оформленных тяжей ― псевдофекалий.
Кровеносная система мидий незамкнутая и состоит из сердца, сети артериальных сосудов,
венозных каналов, лакун и синусов. Сердце делает около 30 сокращений в минуту.
Половой диморфизм не выражен. Определение пола возможно лишь в период
размножения. Самцы отличаются от самок только по цвету зрелой гонады: у самцов ― кремово-
белая, у самок ― оранжевая.
Исследование процессов размножения и раннего онтогенеза мидий имеет первостепенное
значение как для организации рационального промысла, так и для их успешного
культивирования.
В своем развитии мидия проходит через три основные стадии: яйцо, личинка
и собственно особь. Половозрелые особи мидий выметывают яйцеклетки и сперматозоиды
в воду, где и происходит оплодотворение. В среднем один вымет у беломорских мидий длится
около 1 ч [4]. Яйца, имевшие до вымета неправильно-овальную форму, в воде становятся
шарообразными. Первыми начинают вымет самки, хотя Т. А. Матвеева (1948) отмечает, что
находившиеся в ванне самцы первые выпустили сперму, а самки начали вымет лишь через
несколько часов [5].
Плодовитость одной мидии варьирует от 5 до 12 млн яиц, которые при благоприятных
условиях самка может выметать за 15 мин [6]. В. В. Кузнецов и Т. А. Матвеева (1948) отмечали
начало процесса созревания половых продуктов мидий в марте, а готовность самок литоральных
мидий Восточного Мурмана к размножению с мая по октябрь [7]. По их данным, каждая
половозрелая самка может выметать от 100 тыс. до 3 млн яиц в течение этого периода.
Наблюдения за нерестом мидий показали, что вымет половых продуктов происходит не сразу,
а обычно растянут во времени и осуществляется порционно, в несколько сроков, перемежаясь
промежутками покоя [5, 8].
Половозрелости мидии достигают на литорали Баренцева моря при размерах 18–20 мм
в возрасте 5–6 лет, хотя отдельные особи со зрелыми гонадами попадаются и у литоральных
мидий четырехлетнего возраста 13–15 мм длиной [5]. По другим данным, литоральные мидии
Восточного Мурмана достигают длины 13–15 мм в возрасте 2 лет [9], 3 лет в условиях разной
температурно-приливной динамики, 1–2 лет в нижних горизонтах литорали [10]
и при содержании в садках [8, 11]. Половозрелые литоральные мидии губы Зеленецкая Западная
имели длину раковины 17–33 мм [12].
Вклад каждой особи в размножение популяции зависит от ее размеров. Индивидуальная
плодовитость моллюсков увеличивается с увеличением размера животного в кубической
зависимости.
Оседание личинок происходит при длине раковины около 300–350 мкм. Личинка вырастает
до таких размеров за 3–5 недель в зависимости от условий существования популяции.
Оседание личинок на коллекторы в губе Зеленецкая Западная отмечено в конце первой
декады августа, сначала в приповерхностном слое. В середине августа спат покрывал пластины
на всех горизонтах (до 3 м), но его наибольшая плотность отмечена на глубине 1 м (50–85 экз.
на пластину) [13].
В губе Дальнезеленецкая осевшие в августе личинки мидий длиной около 2 мм были
представлены в большом количестве (от 50 до 120 тыс. экз. на 1 м2) на слоевищах водорослей,
но не встречались на илисто-песчаном грунте литорали бухты Дальний Пляж [5].
Первичное оседание личинок происходит обычно вблизи мидиевых поселений,
но в основном на субстрате, не занятом взрослыми моллюсками: на нитчатых водорослях,
гидроидах, раковинах усоногих рачков-балянусов. Личинки, осевшие на нитчатые водоросли,
гидроиды и другие субстраты, проводят на них около месяца и достигают
1–2 мм в длину [2].
Лучшая пища для личинок ― жгутиковые водоросли, кроме того, они потребляют мелкий
свежий макрофитный детрит, бактериопланктон и растворенное органическое вещество. Скорость
фильтрации пищевых частиц у личинок мидий составляет при достаточном кормлении 11.4 мкл/ч-экз.
и уменьшается при очень высоких (более 200 кл/мкл) концентрациях водорослей [2].
Если на первом этапе оседание личинок M. edulis является преимущественно
гидродинамическим, пассивным процессом ("первичное осаждение"), то на конечном этапе
место прикрепления личинок определяется как их активным поиском, так и локальными
условиями биотопа. Особое значение для ориентации личинок в период оседания имеет качество
субстрата. При отсутствии подходящего субстрата личинки задерживают метаморфоз и остаются
в планктоне [2].
Установлено, что в период оседания личинка воспринимает определенные свойства
субстрата, такие как его текстура, цвет, освещенность, пространственное расположение [14].
Субстрат, покрытый бактериально-водорослевой пленкой, наиболее привлекателен для личинок
мидий [15], причем личинка может реагировать не на сам водорослевый субстрат, а на вещества,
выделяемые им в воду [16]. Это обстоятельство учитывается в марикультуре мидии
при определении времени постановки коллекторов. На коллекторах, выставленных перед самым
нерестом, были отмечены лишь единичные особи. А наибольшее количество личинок оседает на
коллекторы, установленные заблаговременно, за 6–9 месяцев до нереста. За это время материал
коллекторов не только покрывается определенным ценозом обрастания, но, очевидно, также
освобождается от значительной доли химических веществ, пропитывающих искусственные
материалы [8].
Шансы молоди на выживание существенно повышаются при осаждении в друзах взрослых
особей своего вида.
Линейный рост моллюсков хорошо аппроксимируется уравнениями Гомперца
и Берталанфи. Уравнение Берталанфи основано на предположении, что скорость роста
определяется разностью между конечным размером животного и его размером в данный момент.
Для сублиторальных мидий Воронки Белого моря это уравнение [17] имело вид:
L = 76.4·(1 – е–0.171·t),
где L ― длина раковины (мм) в возрасте t (годы); е ― основание натурального логарифма.
Кривая роста мидий среднего горизонта литорали губы Зеленецкая Западная [12] также
описана уравнением Берталанфи:
L = 69 (1 – е–0.175·t).
Вместе с тем у мидии наблюдаются большие индивидуальные отклонения в величине
годового прироста начиная уже с первых лет жизни.
Скорость роста моллюсков зависит не только от совокупности абиотических условий,
связанных с положением вида в ареале, но в большой степени от локальных трофических
условий. Сравнительное исследование [11] роста литоральных мидий, помещенных в проточный
аквариум, где в рационе доминировал детрит, и в подвесные садки на акватории губы
Дальнезеленецкая, выявило значимость трофического фактора для роста баренцевоморских
мидий (рис. 2). Хотя в аквариум непрерывно поступала вода из придонного слоя верхней
сублиторали, содержащая большое количество детрита, количество фитопланктона в нем было
обычно в 2 раза меньше, чем в губе. В садках условия питания были лучше, и скорость роста,
соответственно, выше, несмотря на то что среднегодовая температура воды в аквариуме была в
среднем на 1 °С выше, чем в губе. В благоприятных трофических условиях скорость роста
беренцевоморских мидий может быть достаточно высока даже в условиях низких температур
(1.5–2 °С).
Культивирование двустворчатых моллюсков обычно состоит из четырех основных этапов:
1) обследование маточного стада ― естественных или искусственных маточных банок со
зрелыми производителями; 2) получение от них посадочного материала (приплод, молодь или
спат); 3) выращивание его до товарных кондиций; 4) сбор урожая с оставлением его части
для пополнения маточного стада.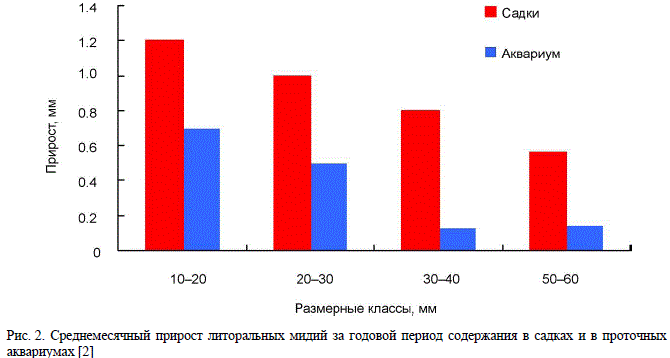
Рис. 2. Среднемесячный прирост литоральных мидий за годовой период содержания в садках и в проточных
аквариумах [2]
Использование мидий происходит по трем основным направлениям: пищевому
(кулинарное и кормовое), фармацевтическому (получение ценных биологически активных
препаратов) и научно-прикладному (мониторинг, биотестирование).
Мировая добыча мидий составляет в среднем около 60–80 тыс. т в год, причем около 50 %
добычи сосредоточено в водах Северной Атлантики, где ведут промысел 13 европейских стран.
Установлено, что состав мяса мидий зависит от условий ее местообитания и сезона.
Содержание влаги варьирует от 45 до 90 %, белка ― от 7 до 11 %, жира ― от 0.5 до 1.6 %,
углеводов ― от 0.2 до 3.5 %, минеральных солей ― от 1 до 2 %. В мясе содержатся в заметном
количестве витамины группы В, витамин D, более 30 микроэлементов. В белке мидий
содержатся все незаменимые аминокислоты, но в большем количестве, чем в яйце. Биологически
активные вещества (БАВ), содержащиеся в теле мидий, содействуют улучшению обмена
веществ, обладают тонизирующим действием [18]. Удельное содержание мягких тканей тела
(мяса) зависит от размеров и возраста мидий и составляет 10–22 % общей массы раковины
для крупных и 16–30 % для мелких моллюсков.
Жирнокислотный состав липидов, выделенных из мяса беломорских мидий,
характеризовался наличием значительных количеств полиеновых кислот. Коэффициент
ненасыщенности жирных кислот превышал в 2 раза величину, установленную для трески.
При сравнительно невысоком содержании водорастворимых витаминов группы В (тиамина
и рибофлавина), что вообще характерно для гидробионтов, уровень токоферолов в теле мидий
оказался на порядок выше, чем в мясе других морских беспозвоночных, например креветок [2].
Поскольку технология культивирования мидий успешно освоена во многих странах мира,
то их марикультура представляет собой хорошую возможность получения дешевого
высококачественного белка и БАВ. Развитие марикультуры моллюсков будет идти, очевидно,
в направлении поликультуры, обеспечивающей бóльшую устойчивость урожаев и полное
комплексное использование ресурсов водоема.
До недавнего времени Баренцево море оценивалось как бесперспективное для разведения
морских беспозвоночных животных [19]. В течение многих лет традиционно считалось,
что только южная зона может быть перспективна для организации мидиевых хозяйств. Однако
к 1975 г. стал известен успешный опыт мидиевых ферм в прибрежных зонах Норвегии
и Шотландии, где в условиях относительно низких температур воды за 14–18 месяцев
в марикультуре были выращены товарные мидии с продуктивностью до 70 т мяса с 1 га [20].
Эксперименты ПИНРО, проводимые на первой опытной мидиевой ферме в 1975–1981 гг., также
показали возможность выращивания на Мурмане мидии съедобной. В условиях полярной
марикультуры мидия достигала товарного размера за трехлетний период [8]. Это более
длительный срок по сравнению с ростом мидий на плантациях Черного моря (где за 1 год
получают до 300 т мяса/га), но при правильной организации мидиевого хозяйства
и бесперебойном подращивании молоди трехлетний цикл выращивания деликатесного мяса
может быть вполне рентабельным [21]. Уже на основании первых практических данных,
полученных на плантации ПИНРО, видно, что опытные фермы на Баренцевом море могут
оказаться более продуктивными, чем это предполагалось ранее. Указывая на теоретическую
возможность подращивания мидии в промышленных масштабах, допускалось, что годовой
валовой сбор в сыром весе мягких тканей мидий может составить около 10–15 т/га поверхности
искусственного субстрата [19].
Обобщенный опыт марикультуры мидий на Мурмане [8] убедительно свидетельствует
о рентабельности мидиевых ферм даже в условиях Заполярья. Установлено, что в условиях
плантации можно получать 10–12 кг мяса с 1 м2 площади за 10 месяцев подращивания,
что при пересчете составит 120 т с 1 га плантации в год [2].
Этот опыт можно успешно применить при утилизации молоди мидий на Баренцевом
и Белом морях, так как первоначальная рентабельность выращивания более крупных мидий
для их кулинарного использования (главным образом как деликатеса) будет, очевидно, невысока.
С учетом продолжительности зимовки, а также ее трудностей при культивировании мидий,
представляется перспективным на первом этапе развития марикультуры на Севере использовать
одно- и двухгодовиков мидий для кормовых целей и технической (и фармацевтической)
переработки.
В настоящее время специалистами ПИНРО разработан способ выращивания мидий
в прибрежной зоне северных морей в бикультуре с лососевыми видами рыб [22].
Коллекторы выполняют в виде полос из долевого полотна длиной 3 м и шириной 30 см,
нижнюю сторону дели снабжают грузом, вес которого достаточен для заглубления коллектора.
В качестве носителей коллекторов используют конструкции плавучей платформы садкового
хозяйства, с внешней и внутренней сторон которой в период с июня по сентябрь коллекторы
для сбора спата мидий закрепляют стационарно на расстоянии около 1 м друг от друга
и погружают на глубину 0.5–1.0 м от водной поверхности. Такая глубина погружения
коллекторов будет оптимальна в любой период культивирования моллюсков [22].
Осевшие на коллекторы мидии культивируют без изменения положения коллекторов в двух-
или трехлетнем цикле в зависимости от поставленных задач по переработке продукции
и по окончании установленного цикла собирают урожай мидий преимущественно в конце мая ―
июне до начала нереста моллюсков, когда в них содержится наибольшее количество качественного
мяса [22].
В качестве груза для заглубления можно использовать достаточно тяжелые предметы,
например камни и иные подручные материалы, которые закрепляют на нижнем конце полосы
делевого полотна.
При этом органические вещества ― отходы жизнедеятельности рыб ― дополнительно
создают среду для развития микрофлоры и фитопланктона, повышая "кормность" среды
для мидий ― этих активных фильтратов, что в конечном счете способствует повышению
продуктивности мидий не только по сравнению с мидиями с природных банок, но и с мидиями,
культивируемыми в монокультуре [22].
Осевшие на коллекторы мидии не подвергаются воздействию отливов, периодически
осушающих литоральные поселения мидий и влекущие за собой соответствующую
прерывистость в поступлении пищи. Мидии на субстратах постоянно находятся в согретом,
хорошо аэрируемом слое воды, толщина которого учтена при определении размеров субстратов-
коллекторов для культивирования моллюсков [22].
В возрасте два года урожай мидий составляет 15 кг, три года ― 16–20 кг с одного
коллектора размером 3.0×0.3 м.
При выращивании мидий на пищевые цели длительность культивирования не превышает
трех лет: в этом возрасте на коллекторах преобладает мидия размером более 50 мм.
Если урожай мидий направлять на медицинские цели, то мидию можно культивировать
два года, в этом возрасте общая биомасса мидий на коллекторе почти одинакова с мидиями
трехлетнего возраста, но количество товарной мидии не превышает 10–15 % [22].
Исландский гребешок Chlamys islandica O. F. Müller, 1776 (рис. 3) ― двустворчатый
моллюск, относящийся к отряду Pectinoida, семейству Pectinidae.
Гребешок широко распространен в Арктике и бореальных водах Северной Атлантики.
У берегов России встречается в Баренцевом и Белом морях на глубинах от 5 до 100 м
на ракушечных и песчаных грунтах, илистых грунтов избегает. Локальные плотные скопления
может образовывать практически на всех глубинах, где обитает. Общая площадь обитания
гребешка в Баренцевом море превышает 40 тыс. км2.
У Мурманского побережья гребешок обитает обычно в диапазоне глубин 5–150 м,
преимущественно на песчаном грунте с примесью ракуши, галечно-гравийного
и крупнообломочного материала, хотя может встречаться начиная с глубин 1–2 м, а также
на каменистых и скальных грунтах [13]. Выделено несколько обособленных поселений,
приуроченных к участкам с неровным дном. Наиболее обширные и плотные поселения
расположены на глубинах менее 100 м в Семиостровской провинции, у о-ва Нокуев, в губе
Савиха, северо-восточнее и восточнее (в Воронке Белого моря) м. Святой Нос и на Южно-
Канинском мелководье (Канинской банке).
В местах обитания динамика придонных вод активная, соленость варьирует от 33.2
до 34.8 ‰, температура ― от 0 до 7.2 °С [23]. Как и у других массовых видов рода Chlamys,
скопления исландского гребешка приурочены к склонам банок и желобов, в районах
гидрологических фронтов [23].
Такое распределение может быть связано как с улучшенным снабжением пищей
(фитопланктоном, детритом), так и с концентрированием личинок в локальных круговоротах
[23]. Оптимальная скорость течения 20–25 см/с: при таких значениях на крупнопесчаных осадках
численность и биомасса достигают максимальных значений 50 экз/м2 и 4 кг/м2 соответственно.
При скорости течения ниже 5 и выше 40 см/с гребешки обнаружены не были [23].
Рис. 3. Исландский гребешок Chlamys islandica
Мягкое тело гребешка заключено в округлую, слабо разностворчатую, равностороннюю
раковину с небольшими выростами у макушки ― ушками. Наружная скульптура створок
представлена радиальными ребрами, дистальная часть которых покрыта двухнедельными
чешуйками роста, образующими достаточно выраженные концентрические кольца. Замковый
край раковины ― прямой без зубов. Лигамент состоит из двух частей: внутренней и наружной.
Внутренняя часть очень хорошо развита и располагается в треугольных ямках под макушками,
наружная имеет вид тонкой пластиночки, соединяющей верхние прямые края створок. Окраска
раковин очень разнообразна и варьирует от коричнево-фиолетовой, ярко-красной или оранжевой
до белой. Верхняя створка, как правило, покрыта темноокрашенными концентрическими
кольцами и немного более выпукла, чем нижняя. Нижняя створка окрашена бледнее и имеет
у основания переднего ушка биссусный вырез, по краям которого располагается несколько зубчиков.
Изнутри раковина выстлана мантией, охватывающей с боков туловище моллюска.
Мантийные лопасти, снабженные по краям щупальцами и глазами, не срастаются на брюшной
стороне и образуют так называемый парус, закрывающий мантийную полость при плавании,
которое осуществляется по принципу реактивного движения, как правило, макушкой назад.
Прикрепление гребешков к субстрату происходит с помощью тонких клейких нитей,
выделяемых биссусной железой, расположенной в ноге.
Нога развита слабо, мышц-элеваторов нет, есть только один ретрактор, асимметрично
прикрепляющийся к левой створке. Единственный мускул-замыкатель (аддуктор) разделен
на два неравных отдела: большой ― передний, состоящий из поперечнополосатых мышечных
волокон, и задний, слагающийся из гладких волокон. Кроме вышеперечисленных, у гребешков
имеются еще небольшие мышцы, которые поддерживают околосердечную сумку, фиксируют
жабры и выполняют другие функции.
Щелевидное ротовое отверстие расположено на передней стороне тела. Пищей моллюскам служит
фито-, зоо-, бактериопланктон и взвешенный детрит, приносимые с водой в мантийную полость.
На Мурманском побережье половое созревание гребешка начинается в 3 года и
заканчивается к 6 годам: в этом возрасте все особи являются половозрелыми. У западной
Гренландии этот процесс происходит с 4 до 9 лет при высоте раковины 30–55 мм. В Норвегии ―
с 5 до 7 лет при 40–50 мм. В раннем возрасте моллюски созревают преимущественно как самцы,
но к 5 годам соотношение между полами выравнивается и становится близким к 1. Форма
раковины самок и самцов не отличается. Самки отличаются от самцов цветом гонады: у самок
она оранжевого цвета, у самцов ― белого. Существует предположение, что вследствие
относительно более высоких энергетических затрат на размножение выживаемость самцов ниже,
чем самок, и у старших возрастов это может приводить к снижению их доли
в соотношении полов [24]. Нерест происходит с апреля по сентябрь, и его конкретные сроки
определяются, по одним данным [23], накопленной за год температурой в 380 градусодней,
по другим [25] ― началом развития фитопланктона.
В Баренцевом море, на западе у о-ва Медвежий, нерест происходит в апреле ― мае и по
мере продвижения на восток смещается на более поздние сроки: у западного побережья п-ова
Канин он происходит в июне ― июле. Плодовитость связана с высотой раковины
аллометрической зависимостью (показатель степени b = 5.2) и в среднем равна 5.2 млн яиц [23].
Оплодотворение наружное. Пелагический период личинки длится 9–10 недель [26].
Оседание происходит преимущественно на твердые субстраты, куда личинки
прикрепляются биссусом. Один из наиболее предпочтительных субстратов ― мертвый перисарк
гидроидов рода Tubularia: личинки гребешка предпочитают перисарк живым гидроидам,
красным водорослям и различным искусственным субстратам. Более того, "чистый" перисарк
более предпочтителен, чем перисарк, покрытый бактериальной пленкой. Вероятно, основную
роль в привлечении молоди играет белковый и/или полисахаридный компонент мертвого
гидроида. Так же как и у мидии, осевшая молодь гребешка способна открепляться, некоторое
время пребывать в толще воды, а затем заново прикрепляться. При этом отмечено, что повторное
прикрепление обычно осуществляется к тому же субстрату, на который первоначально осела
личинка. На коллекторы оседают преимущественно личинки, хотя через 3–4 месяца вторично
осевшая молодь и обнаруживается в небольшом количестве [13].
Ярко выраженная сезонность роста, свойственная видам, населяющим высокие широты,
позволяет сравнительно легко определять возраст по наружным кольцам и/или по меткам
на обызвествленном основании лигамента. Применение обоих методов в комбинации с прямыми
наблюдениями за осевшей молодью позволили оценить темпы роста и возрастной состав
поселений в разных частях ареала.
На коллекторах за 1 год молодь дорастает до 7 мм в Норвегии и до 9.5 мм в Исландии.
За 2–3 года в Исландии гребешки в подвесных садках дорастают до 25 и 40 мм соответственно.
В естественных поселениях у западной Гренландии средний возраст моллюсков составляет
21 год при высоте раковины 87 мм. В Исландии максимальный возраст также составляет 20 лет,
а максимальный размер достигает 140 мм. На Мурманском побережье в Семиостровском
поселении максимальный возраст ― 18 лет, до 90 мм гребешки дорастают в среднем за 8 лет [13].
Наиболее высокая скорость роста отмечена в районах, где соотношение между
взвешенным неорганическим материалом и органикой минимально. Так же как и мидии,
в толще воды (подвесная культура) гребешки растут лучше, чем на дне, и это объясняется более
высокой долей фитопланктона в рационе и, соответственно, более низкой долей детрита. Так,
в период весеннего цветения зарегистрированы и максимальные темпы роста. Возможно,
на скорость роста влияет и внутривидовая конкуренция, так как даже в подвесной культуре
лучше росли особи в садках, расположенных не над гребешковой банкой, а в стороне от нее.
Смертность наиболее высока на фазе личиночного развития (выживает одна личинка
из 2.5 млн). В первый год жизни коэффициент естественной убыли равен 1.13 год–1. Начиная
с третьего года и до девяти лет смертность фактически постоянна и составляет 0.12 год–1,
и только у моллюсков старше девяти лет она увеличивается до 0.2 год–1. Вследствие сильного
пресса хищников (морских звезд, крабов, донных рыб) молодь обитает преимущественно
в убежищах: под ракушками, между валунами, в щелях [13]. Основные промысловые скопления
гребешка расположены у мыса Святой Нос (южнее 69°с. ш.) преимущественно на глубине
70–100 м и в Воронке Белого моря преимущественно на глубине 40–60 м. В период
2000–2009 гг. наблюдалась тенденция уменьшения промыслового запаса гребешка
в большинстве скоплений Баренцева моря (рис. 4).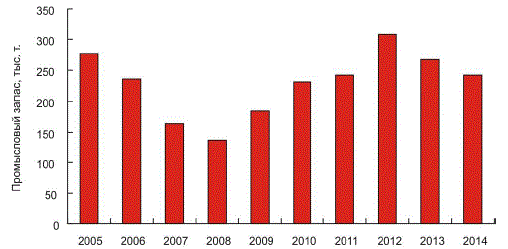
Рис. 4. Динамика промыслового запаса исландского гребешка в Баренцевом море в 2005–2014 гг. [27]
Наибольшее снижение запаса отмечалось на участке Святоносского поселения,
располагающегося в исключительной экономической зоне РФ [27]. Вероятными причинами
этого являлись негативное влияние промысла гребешка и донных видов рыб, отсутствие
достаточного пополнения промыслового запаса, а также повышенная естественная смертность
от болезней. В 2010–2014 гг. промысловые скопления гребешка сохранились только в зоне
территориальных вод РФ и то лишь на той части акватории, где наблюдалось достаточное
пополнение промыслового запаса [27].
В период промысла гребешка в Баренцевом море максимальные уловы наблюдались
в 1997–1999 гг. ― до 14 тыс. т в год, но в последние 10 лет улов значительно снизился
и не превышал 3.1 тыс. т. (рис. 5).
Основным фактором, определяющим уменьшение вылова, было сокращение площади
промысловых участков с высокими плотностями гребешка, что привело к снижению
промысловых усилий и количества судов на промысле (с 5 до 1 ед.). В соответствии
с рекомендациями ПИНРО, в 2009–2010 гг. промысел не проводился. С 2011 г. добыча
возобновилась, но вылов, как правило, не превышал 0.5 тыс. т. (рис. 5). В 2014 г. (по данным
на 1 декабря) было выловлено 817.8 т, средняя производительность промысла достигала 19.5 т
за судо-сутки [27].
Культивирование гребешков имеет достаточно давнюю традицию, особенно в Японии,
где успешно выращивают приморского гребешка Mizuhopecten yessoensis (Jay). В качестве
основного способа выращивания этого вида в российских водах специалистами ТИНРО выбран
экстенсивный метод, который заключался в сборе молоди на коллекторы и подращивании
в садках. В ряде обобщающих работ [28, 29] проведен детальный анализ состояния
марикультуры приморского гребешка на Дальнем Востоке. Процесс культивирования гребешков
в этом районе был разделен на два этапа. На первом этапе проводят сбор оседающей молоди
моллюсков и размещение ее на коллекторах, изготовленных из искусственных и естественных
субстратов. Время сбора спата и сроки его подращивания в садках выбирают с учетом процессов
природного гаметогенеза, что необходимо для прогнозирования объемов оседания личинок.
Чаще всего собранный спат размещают на грунте или в садках близ водорослевых плантаций,
где в течение года осуществляется промежуточное выращивание молоди.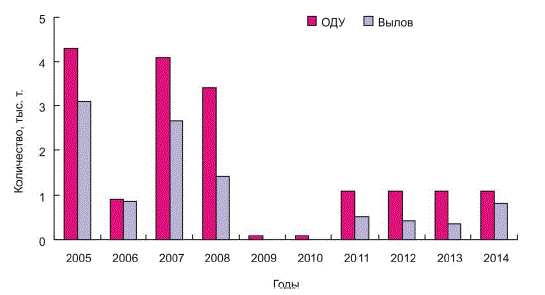
Рис. 5. Общий допустимый улов (ОДУ) и вылов исландского гребешка в Баренцевом море в 2005–2014 гг. [27]
Время выставления коллекторов в море определяют исходя из информации о плотности
и качественном составе личинок гребешка в планктонных пробах.
После достижения молодью гребешка размеров 8–10 мм (обычно в августе) гирлянды
коллекторов снимают с установок и разбирают на специальной понтон-площадке.
Для выращивания в подвесной культуре отсортированную молодь помещают в садки (площадь
дна садка 0.12 м2) с плотностью 20 экз/садок, тогда как для последующего перевода в донную
культуру ― с плотностью 250 экз/садок. Заполненные садки собирали в гирлянды, состоящие
из 10 однотипных элементов, и вновь размещали на горизонтальных канатах установок. Осенью
установки с садками притапливали на 1.5–2 м от поверхности воды.
В мае следующего календарного года моллюсков, предназначенных для выращивания
в подвесной культуре, пересаживали в новые садки, снижая плотность до 10 экз/садок.
При такой плотности их содержали на установках до конца технологического цикла.
В это же время молодь гребешка, содержавшуюся с плотностью 250 экз/садок,
освобождали из садков и помещали в транспортные емкости, в которых моллюсков доставляли
морем к местам отсадки на грунт. Рекомендованная начальная плотность при выращивании
гребешка в донной культуре составляла 10–15 экз/м2.
Продолжительность технологического цикла при выращивании гребешка в подвесной
культуре была три, а для донной культуры ― четыре года. Расчетная урожайность в эквиваленте
прижизненной массы достигала 26 и 8 т/га соответственно.
За последние 30 лет предложенные технологические схемы не претерпели существенных
изменений, а объемы выращивания гребешка в Приморье так и не достигли промышленных.
Эксперименты и полевые наблюдения показали, что выращивание гребешка в бикультуре
с ламинарией позволяет увеличить скорость его роста и индекс кондиции. Во время второго
этапа уже подросших моллюсков переводят в специальные садки, где они достигают товарных
размеров. Иногда доращивание гребешков проводят на грунте, но в таких условиях моллюски
растут медленнее. В ходе экспериментов по подращиванию гребешков выяснилось,
что личиночное пополнение имеет нерегулярный характер и его интенсивность может
варьировать в широких пределах [30]. Причиной этого может быть разрушение поселений
гребешка, перемещающихся вместе с песком под воздействием штормового волнения
в незащищенных рельефом участках дна. Кроме того, после двух лет подращивания в садках
отмечается резкое уменьшение численности расселенной на грунте молоди гребешка
за счет естественной убыли животных. Все это очень усложняет расчет усилий, необходимых
для организации производства. Расчеты показали, что донное выращивание гребешков менее
рентабельно, чем содержание особей в садках до товарного размера, т. е. 5–7 см.
Спорадичность и непредсказуемость объемов пополнения популяции молодью потребовало
научных разработок по получению личинок и подращиванию молоди гребешка в контролируемых
заводских условиях. Изучены способы регуляции гаметогенеза, стимуляции нереста гребешков [31],
пищевые потребности и энергозатраты [32]. Разработаны также необходимые смеси и рационы
питания для личинок гребешка и спата [33]. С целью интенсификации оседания личинок в
специальных резервуарах были опробованы аттрактанты, оказавшиеся очень эффективными.
В условиях изменившихся в последнее десятилетие социально-экономических обстоятельств,
особую актуальность приобретает не принципиальная возможность демонстрации того или иного
способа выращивания гребешка, а экономическая целесообразность реализации таких
технологических схем культивирования этих моллюсков, которые обеспечивают промышленные
масштабы и приемлемую рентабельность хозяйств марикультуры [28].
Для выявления возможности и целесообразности культивирования исландского гребешка
учеными ММБИ осуществлена серия экспериментов по подращиванию 3–4-летней молоди
моллюсков этого вида в губе Дальнезеленецкая Восточного Мурмана [23, 34]. Первоначально
проводились наблюдения за выживаемостью гребешков, свободно подвешенных
на вертикальных гирляндах в толще воды. Каждая гирлянда представляла собой капроновый
шнур длиной 10 м, от которого через равные промежутки отходили 15 поводков из прочной
лесы. Свободный конец поводков продевался сквозь отверстие, высверленное в переднем ушке
раковины моллюска, и завязывался узлом, не препятствующим раскрытию створок. Семь таких
гирлянд с подвешенными к ним гребешками были закреплены на одном плавучем основании
и размещены в защищенном от шторма месте глубиной 15 м (рис. 6, А) вблизи о-ва Немецкий
(губа Дальнезеленецкая).
Месячная экспозиция моллюсков на установке в ноябре 1978 г. показала, что они хорошо
перенесли операцию по высверливанию отверстия в ушке и внешне ничем не отличались
от интактных особей с неповрежденной раковиной. Внутренняя поверхность створок, частично
оголившаяся в момент перфорации вследствие сокращения паллиальной мускулатуры, к концу
месяца полностью закрылась мантийной складкой. Гибели гребешков в течение всего периода
наблюдений не отмечено.
В июле 1979 г. между островами Сухим и Жилым (в той же губе) на глубине 10 м была
поставлена новая усовершенствованная установка, несущая 10 гирлянд и 20 конических садков
(рис. 6, Б). В садках диаметром 40 см находилось 200 экз. моллюсков, на гирляндах длиной
8 м ― 160 экз. Перед началом подращивания все гребешки были промерены, а помещенные в
садки, в дополнение к этому, помечены стоматологическим инструментом. Наблюдения
за подращиваемыми моллюсками в течение первых двух месяцев проводились ежедекадно,
а в дальнейшем ― ежемесячно. Регулярному осмотру на воздухе подвергались только особи
опытной группы, составляющей ровно половину всей совокупности. Контрольная группа
находилась в воде вплоть до окончания эксперимента в июне 1980 г. [34].
В первый месяц подращивания на гирляндах отмечена массовая гибель моллюсков,
подвешенных в слое 0.5–1 м. Вероятно, это объясняется проникновением сквозь тонкий слой
воды ультрафиолетового излучения, угнетающе действующего на многих гидробионтов. К концу
третьей недели раковины гребешков покрылись густым войлоком нитчатых водорослей.
На внутреннем участке створок, оголенном на ширину 5–8 мм в результате сокращения
травмированной мантии, появилась масса одноклеточных водорослей. В дальнейшем по этой
причине у большинства особей наблюдался патологический рост раковины.
Активная кальцификация не затрагивала покрытую водорослями внутреннюю поверхность
створок, и новые слои роста формировались в виде козырька над пораженной зоной. В целом
годовой прирост на гирляндах в опытной и контрольной сериях статистически не различался,
от глубины подращивания не зависел и составил в среднем 5±4 мм. Суммарная смертность
моллюсков данной совокупности к концу срока наблюдений достигла 34 %, причем на долю
опытной группы пришлось 38 %, а на долю контрольной ― 30 %.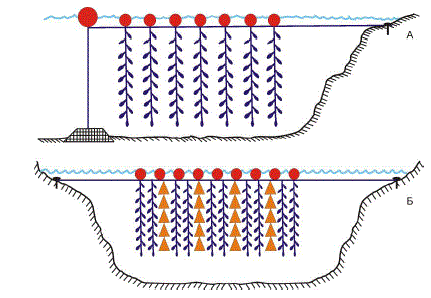
Рис. 6. Схема установок, применявшихся для подращивания гребешков в 1978 (А) и 1979 гг. (Б) [34]
В садках в первые дни эксперимента такая гибель гребешка, как на гирляндах, отмечена
не была. Быстрое обрастание конуса конструкции нитчаткой послужило для находившихся
внутри моллюсков надежной защитой от губительного воздействия ультрафиолетового
излучения. Рост особей с нетравмированной мантией происходил в виде правильной
кальцификации по краю раковины. Если на гирляндах момент начала роста установить было
невозможно, то в садках увеличение размеров моллюсков за счет формирования новых слоев
раковины стало заметным на четвертой неделе эксперимента. Годовой прирост в контрольной
серии к концу периода наблюдений достиг 15±3 мм и статистически несколько отличался
от прироста в опытной серии, составившего 11±3 мм. Суммарная элиминация за время
подращивания оказалась вдвое ниже естественного уровня смертности и составила всего 6 %.
Так же как и на гирляндах, в опытной группе она была выше (7 %), чем в контрольной [34].
Интересным результатом экспериментов явилось то, что у всех моллюсков на гирляндах
и в садках новые, сформировавшиеся за время подращивания слои раковины приобрели
красивую красно-пурпурную окраску. Произошедшие изменения изначальной расцветки створок
(серо-белой, фиолетово-розовой и коричневой), вероятно, были связаны с повышенным
содержанием каротиновых пигментов в сестоне поверхностных слоев воды. В случае
подтверждения полученных нами результатов представляется возможным практическое
использование данного эффекта при культивировании гребешков не только для получения
пищевой и кормовой продукции, но и с целью использования раковин моллюсков
для производства декоративных изделий и сувениров. Подводя итог результатам эксперимента,
необходимо отметить, что подращивание гребешков в садках выгодно отличается
от подращивания на гирляндах за счет более высоких темпов роста и пониженной смертности [34].
На основе данных эксперимента и анализа эколого-физиологических характеристик
популяций исландского гребешка оценена возможность промышленного культивирования
гребешков в Баренцевом море [35]. Организация хозяйства по культивированию исландского
гребешка на Баренцевом море рассмотрена в трех вариантах:
1) выращивание на коллекторе от личинки (спата) до года с последующим расселением в садки;
2) выращивание на коллекторе от личинки (спата) до года с последующим расселением на грунт;
3) сбор 2–3-летней молоди водолазами (промысловыми судами) с последующим
подращиванием в садках до товарных размеров.
Варианты различаются периодами (сроками) выращивания гребешков до товарной
продукции: 1-й ― 5 лет, 2-й― 7 лет, 3-й ― 3 года.
На первом этапе по первому и второму вариантам осуществляется сбор спата
на коллекторы рамной установки с последующим выращиванием до года. При нормативном
сборе спата на одном коллекторе 300 экз. и 20 % отхода для выращивания остается 250 экз.
Общий объем товарного спата на одной установке ― 5.3 млн экз. (250×21210), где
21210 ― число коллекторов на одной установке. Через год спат по первому варианту
пересаживается в садки по 250 экз. (первый вариант) или расселяется на грунте (второй вариант).
На следующем этапе первого варианта двухгодовиков пересаживают по 20 экз. в садок.
При выживаемости 90 % моллюсков второго года выращивания для пересадки потребуется
11 установок. На последнем этапе плотность посадки трехгодовиков ― 10 экз/садок.
При выживаемости 95 % число установок увеличивается до 22.
К концу пятого года численность выращенных гребешков составит 4486 млн экз., товарная
масса сырца ― 408.1 т, масса мяса мускула ― 44.8 т. Аналогичные результаты предполагаются
для шестого года и по второму варианту [34].
Беспрерывная замкнутая схема выращивания в первом варианте осуществляется
при эксплуатации установок по сбору спата и подращиванию двухгодовиков, 11 установок
для трехгодовиков и 44 ― для завершающего двухлетнего периода. В целом предполагается
использование 57 установок.
В третьем варианте планируется собранных водолазами 2–3-летних гребешков выращивать
с плотностью посадки 20 экз/садок и пересадкой через год по схеме первого варианта.
Для обеспечения равных объемов с первым вариантом необходимо 55 (11+44) установок.
В зависимости от глубины моря (10–35 м) время работы водолаза под водой колеблется
от 1 до 2 ч. В среднем принимаем 1.5 ч. Одновременно работают два водолаза, в течение
смены ― три пары. При двухсменной работе общее время сбора гребешков двумя водолазами
составит 9 (1.5×3×2) ч/сут.
Расчеты показывают, что прибыльность обеспечивает второй вариант, наиболее убыточен
третий, рентабельность которого не обеспечивается даже заменой водолазных работ сбором
молоди из приловов в ходе промысла моллюсков судами. Проблематичным при реализации
второго варианта является достаточная обеспеченность водолазными работами. Это же, помимо
относительно высокой капиталоемкости, относится и к первому варианту [34].
На Северном бассейне при переработке используется лишь мускул гребешков. Это снижает
возможности повышения цен на сырец. Однако в мировой практике находят применение
и другие части тела моллюсков (гонады, мантия). Более высокая степень утилизации
(при переработке) исландского гребешка позволит повысить в 1.5–2 раза цены на сырец. В этих
условиях культивирование гребешков в Баренцевом море станет экономически приемлемым
даже по первому варианту [34]. Более рациональной будет организация умеренного вылова
гребешков в сочетании с недорогостоящими мероприятиями по пополнению их естественных
ресурсов путем сбора на коллекторы оседающей молоди и расселения ее по подходящей акватории.
Основные проблемы в организации аквакультуры в Баренцевом море, как и в других
водоемах РФ, связаны со следующими факторами [36]:
• на первоначальном этапе развития предприятий аквакультуры требуются крупные
денежные средства и длительный цикл выращивания объектов;
• отсутствует информация о пригодности акватории для выращивания объектов
марикультуры, что влечет за собой дополнительные расходы средств и времени на обследование
участка и разработку биологического обоснования;
• подорваны естественные запасы гидробионтов, которые не позволяют новым
предприятиям осуществлять сбор спата (молоди) в должном объеме;
• недостаточно эффективна финансовая поддержка, устарели производственные мощности
и материально-техническая база, слабо развит механизм страхования рисков, отсутствует
инновационная деятельность.
Подводя итог, отметим, что потенциальные возможности использования двустворчатых
моллюсков в аквакультуре очень велики. Это обусловлено особенностями их биологии, среди которых
можно выделить следующие важнейшие черты: фильтрационный тип питания, близость к начальным
звеньям трофической цепи; относительно короткий жизненный цикл; относительно быстрый рост во
всей толще воды, где расположены гидробиотехнические сооружения, до достижения товарных
размеров; устойчивость взрослых моллюсков к колебаниям солености и температуры воды (для
мидий), высокая плодовитость, высокая морфологическая вариабельность в естественных и
искусственных местообитаниях. В настоящее время накоплен достаточный объем информации и
созданы необходимые наработки для организации марикультуры мидий и исландского гребешка в
Баренцевом море. Главным препятствием является относительно долгий период окупаемости
финансовых вложений при организации акваферм на Кольском п-ове.
Полярная аквакультура
МОРСКИЕ БИОЛОГИЧЕСКИЕ РЕСУРСЫ СЕВЕРНОГО БАССЕЙНА: ПРОМЫСЕЛ, ПОТЕНЦИАЛ И ПРОБЛЕМЫ РЕГУЛИРОВАНИЯ



