АНАЛИЗ ТИПОВ ЖИЗНЕННЫХ СТРАТЕГИЙ МОРСКИХ ЕЖЕЙ КАК МЕТОД ОЦЕНКИ ПЕРСПЕКТИВНОСТИ ИХ ИСПОЛЬЗОВАНИЯ В АКВАКУЛЬТУРЕ
Функциональные характеристики морских ежей могут быть использованы для оценки пригодности видов для аквакультуры. Наиболее важные функции могут быть разделены на две основных категории: продукция биомассы и поддержание жизнедеятельности (восстановление поврежденных и утраченных частей тела). Виды, обитающие в биотопах, которые или являются нестабильными по ряду физических факторов, или с высокой степенью хищничества и с высокой доступностью (обилием) пищи, должны иметь высокую продукцию и низкие траты на поддержание своей жизнедеятельности. Высокая продукция и умеренные траты на поддержание жизнедеятельности, а также высокий потенциал к фенотипической пластичности, должны встречаться у видов, обитающих в стабильных биотопах с низким прессом хищничества и высокой доступностью пищи. Виды, обитающие в стабильных биотопах или с низкой степенью хищничества и с низкой доступностью пищи, должны иметь низкую продукцию и высокие
Еаты на поддержание жизнедеятельности. Рассматриваются морские ежи родов Tripneustes, ongylocentrotus и Arbacia, как реализующие эти типы стратегий. При использовании продукции как основного критерия можно предположить, что виды в первой категории наиболее перспективны для использования в аквакультуре. Предлагается схема вариантов реализации типов жизненных стратегий видами рода Strongylocentrotus в различныхусловиях. Исторически использование некоторых видов морских ежей в качестве промысловых могло быть связано с высокой степенью их доступности и вкусовыми качествами и не обязательно указывает их пригодность для аквакультуры. Выявление функциональных особенностей видов может помочь в оценке перспективы использования морских ежей в аквакультуре.
Несмотря на большое количество видов морских стран, в качестве промысловых употребляются ежей, обитающих в прибрежных водах разных лишь некоторые. На первый взгляд, не всегда очевидно, какие критерии использовались потребителями при выборе видов. В частности, при доступности и большом обилии прибрежного морского ежа Tetrapygus niger в Чили (Vasquez, Buschmann,
1997), он практически не используется в пищу местным населением. Основной причиной игнорирования этого вида, как оказалось, является отсутствие вкусовой привлекательности его гонад. Аналогичная ситуация наблюдается с морским ежом Arbacia lixula из того же семейства. Этот вид употребляется в пищу в средиземноморских странах только в некоторых районах, в то время как Paracentrotus lividus широко промышляется в данном регионе (Boudouresque, 1990).
Икра морских ежей сем. Arbacia, помимо своей вкусовой непривлекательности, имеет темнофиолетовый цвет, в отличие от обычно желтой или оранжевой икры традиционных промысловых видов морских ежей, таких как P lividus и стронги-лоцентротиды.
Необычный цвет может сигнализировать о наличии продуктов вторичного метаболизма, предупреждающих потенциальных хищников о несъедобности или даже ядовитости потенциальной жертвы (Harvey, Greenwood, 1978). Характер подобного предупреждения является важным и для выметанных яиц, свободно плавающих в толще воды и уязвимых для хищников. Почему виды сем. Arbacia производят дополнительные энергетические траты, вырабатывая вторичные метаболиты, в то время как P. lividus и виды рода Strongylocentrotus не делают этого? Может ли данный специфический вид метаболизма быть связан с типом стратегии? И, если связан, может ли анализ стратегий быть важным в оценке перспективности видов морских ежей для использования в аквакультуре? Имеют ли стратегии свою основу в энергетических потребностях и тратах, которые могут быть использованы в понимании функциональных особенностей, необходимых для оценки их потенциала? В данной статье предлагается использовать некоторые основные видовые характеристики, связанные со стратегиями видов, полагая, что они могут быть вполне полезны для данной цели.
МАТЕРИАЛ И МЕТОДИКА
Основной материал был собран в экспедициях Камчатского отдела Института биологии моря ДВО РАН, Тихоокеанского института географии ДВО РАН, Камчатского отдела природопользования Тихоокеанского института географии ДВО РАН, Камчатского отдела морской биологии и биотехнологии Тихоокеанского океанологического института ДВО РАН, Института биологии моря ДВО
РАН, на побережье Восточной Камчатки от м. Лопатка до б. Оссора, на Северных Курильских островах: о. Атласова, о. Шумшу, о. Пара-мушир, в Беринговом море: о. Беринга, о. Медный, зал. Олюторский, побережье Корякского нагорья, в Амурском заливе, заливе Посьет (Японское море), на о. Сахалин, с 1979 по 1991 гг.
Помимо собственных сборов, обработаны материалы ряда экспедиций из коллекций Зоологического института РАН, Института биологии моря ДВО РАН и Института океанологии РАН.
Всего за период исследования были просмотрены 29 293 образца с 916 станций. Из них обработано по видам: S. pallidus — 9627 образцов, S. droeba-chiensis — 5130, S. polyacanthus — 10799, S. intermedius — 2376, S. nudus — 1370 образцов.
При работе на глубине до 30-35 м использовали легководолазное снаряжение. Гидробиологические разрезы закладывали перпендикулярно береговой линии. Пробы отбирали с интервалом 2-5 м по глубине, проводя тотальный сбор животных, в пределах мерной рамки площадью 0,25 или 1 м2. Рамки перекладывали на дне от трех до двадцати пяти раз, в зависимости от плотности поселений морских ежей. Минимальное суммарное число учтенных особей составляло 30-35. На глубине 35-250 м пробы отбирали гребешковой драгой шириной захвата 3,5 м и дночерпателем «0кеан-0,25». Выполнено 605 гидробиологических станций.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Энергетический баланс
Согласно второму закону термодинамики, живые системы должны использовать энергию для поддержания своей жизнедеятельности во времени (Bertalanffy, 1952). Как открытые системы, организмы делают это, получая энергию из окружающей среды и размещая ее в различные виды деятельности. Связь между полученной энергией и ее распределением выражена в уравнении баланса энергии (Ricker, 1968):
C - F = A = R + U + Pr + Ps, где C — потребление, F — фекалии, A — адсорбция, R — дыхание, U — выделение, Pr — репродуктивная продукция, и Ps — соматическая продукция. Энергия, связанная с дыханием, тратится на анаболизм и поддержание жизнедеятельности. Ps используется на рост и восстановление скелетных структур для защиты тела. Pr и Ps вместе составляют общую продукцию или ассимиляцию. При расчете энергетического баланса используются единицы энергии (джоули). Другие единицы (масса) могут быть использованы для других целей, но
только единицы энергии позволяют проводить вычисление относительного, не абсолютного, распределения энергии на использование в различных функциях, рассмотрение которых является существенным для установления типов стратегий (Calow, Townsend, 1981).
Известны только немногочисленные попытки составления энергетического баланса для морских ежей, и все они не могут считаться полными. Например, в бюджете энергии для Strongylocentrotus intermedius, предложенном Фуджи (Fuji, 1967), недостает рассмотрения аспектов дыхания, а в энергетическом балансе S. droebachiensis М.В. Проппа (1977) явно не хватает аспектов потребления.
Типы стратегий
Выбор стратегии может быть связан с энергетическим балансом, лежащим в основе концепции стратегии (Ebert, 1982). В энергетической системе организма существуют различные варианты переключения путей энергетических трат: на соматический рост, гонадную продукцию и на выживание, причем при поступлении ее на поддержание одной функции энергия становится не доступна для других функций (Calow, 1984).
Жизненные стратегии отражают, как эти характеристики изменяются, чтобы увеличить соответствие окружающей среде (Stearns, 1992). Взаимодействие этих характеристик четко выражено в уравнении Эулер-Лотка (Calow, 1964):
1 = Ze-rtsn,
где r — параметр Мальтуса (скорость экспоненциального роста), t — время до наступления поло-возрелости, функция соматической продукции; s — выживание, функция поддержания жизнедеятельности (восстановление поврежденных и утраченных частей тела) и защиты организма; и n — плодовитость, функция гонадной продукции. Соответствие организма окружающей среде увеличивается при увеличении s и n и при уменьшении t.
Грайм (Grime, 1977) связал стратегии жизни с двумя внешними факторами, которые ограничивают энергию индивидуума. Первый — это воздействие стресса, который ограничивает продукцию биомассы. Он включает в себя слабую доступность пищи, низкую способность к питанию и суб-оптимальные абиотические условия. Другой фактор — нарушение, приводящее к потере биомассы. Обычно это воздействие, определяемое хищничеством или влиянием летальных абиотических факторов. Виды, которые испытывают высокую степень нарушения — рудеральные виды, очевидно, будут иметь низкую степень выживаемости s, высокую скорость роста (незначительное время до
наступления половозрелости t) и высокую плодовитость n. Эти виды расходуют незначительное количество энергии на поддержание и защиту тела, поскольку большая часть энергии используется на рост и воспроизводство. Виды, которые испытывают высокую степень стресса, стресс-толерант-ные виды, очевидно, имеют высокую степень выживаемости, низкую плодовитость и низкую скорость роста. Эти виды расходуют много энергии на поддержание жизнедеятельности или защиту, поскольку она не используется на рост и воспроизводство. Это увеличивает вероятность выживания этих видов длительное время, достаточное для успешного воспроизводства. Виды, которые испытывают незначительную степень нарушения и стресса, виды-конкуренты, используют энергию для обеспечения всех функций.
Эти три первичных типа стратегий встречаются при экстремальных уровнях стресса и нарушения. Очевидно, что вторичные типы стратегий реализуются видами при промежуточном уровне внешних факторов.
Типы стратегий и перспективность видов морских ежей для аквакультуры
Если в качестве основного критерия использовать уровень продукции, то виды морских ежей, наиболее пригодные для аквакультуры, должны иметь самое короткое время до наступления поло-возрелости (высокую скорость роста), самую высокую плодовитость и самую низкую степень выживаемости. Они не должны расходовать много энергии на поддержание жизнедеятельности или защиту тела. С другой стороны, виды, вкладывающие энергию в поддержание жизнедеятельности, оказываются способны лучше противостоять стрессовому воздействию. Д. Лоуренс (Lawrence, 1990) определил специфические типы жизненных стратегий различных видов морских ежей. Возникает вопрос, можно ли использовать эти характеристики для оценки перспективности видов морских ежей для аквакультуры?
Выбор определенного вида для использования в аквакультуре может диктоваться двумя требованиями — доступностью вида или соответствием его набору специфических критериев. Доступность вида является весьма существенным доводом, но пример с Tetrapygus niger показывает, что этого не достаточно. Очевидно, что различные функциональные характеристики связаны с различными типами стратегий видов морских ежей (табл. 1). Виды должны иметь набор определенных характеристик, присущих определенному типу реализуемой стратегии. К примеру, такие виды как
Таблица 1. Предполагаемые функциональные характеристики, связанные с различными типами стратегий видов морских ежей
Tripneustes ventricosus, обитающие в биотопах, подверженных большому уровню нарушений, но с высоким обилием пищи, очевидно, будут иметь высокую скорость роста, высокие репродуктивные усилия, высокий уровень метаболизма, незначительные энергетические траты на поддержание жизнедеятельности и короткую продолжительность жизни. Следует рассмотреть, действительно ли эти характеристики свидетельствуют о пригодности видов морских ежей для аквакультуры.
Скорость роста
Эберт (Ebert, 1975, 1982) разделил виды морских ежей в соответствии с их темпом роста, делая вывод, что такие виды, как Strongylocentrotus droebachiensis растут медленнее, чем Tripneustes ventricosus и Lytechinus variegatus. Сравнение видов может быть сделано лучше всего, если особи питаются одной пищей при прочих равных условиях. Молодь Echinometra mathaei, Diadema setosum и Tripneustes gratilla, при содержании в одном аквариуме, кормили водорослями. Результаты показали существенную разницу в темпах роста (табл. 2). Размер панциря особей, достигших возраста один год, также указывает на значительную видоспецифичную вариабельность в скорости роста (табл. 3). Определение темпа роста молоди является важным моментом, поскольку этот показатель связан с временем наступления половозрелости (табл. 4).
Хотя результаты этих исследований весьма показательны в различиях темпов роста между
Таблица 2. Рост (сырая масса тела, мг) морских ежей, содержавшихся в аквариуме г. Эйлат, Израиль (Lawrence, 2007)
видами, они не дают информации о продукции в единицах энергии. Так, измерение темпов роста у видов, имеющих массивную стенку тела и содержащую большое количество карбоната кальция, даст завышенные результаты по продукции; кроме того, увеличение объема имеет следствием увеличение общей массы тела, связанное, однако, с увеличением количества целомической жидкости, а не с продукцией.
Тем не менее, приведенные данные свидетельствуют о значительных функциональных различиях, которые, как предполагается, непосредственно связаны с различными типами стратегий.
Продолжительность жизни
Т. Эберт (Ebert, 1975, 1982) обнаружил различия в продолжительности жизни видов морских ежей и общую отрицательную корреляцию со скоростью их роста. Используя литературные данные, этот автор (Ebert, 1975) показал большую продолжительность
Таблица 3. Размер (диаметр панциря, мм) различных видов морских ежей в возрасте 1 год
Виды Размер, мм Автор
Пор. Diadematoida
Сем. Diadematidae
Diadema antillarum 40 Lewis, 1966
Diadema setosum* 45 Drummond, 1993
Пор. Phymosomatidae
Сем. Stomechinidae
Stomopneustes variolaris 15 Drummond, 1993
Пор. Temnopleuroida
Сем. Toxopneustidae
Lytechmus variegatus* 50 Moore et al., 1963
« 50 Allain, 1975
« 50-70 Oliver, 1987
Tripneustes gratilla* 50 Shokita et al., 1991
« 70 Dafni, 1992
« 40 Maharavo, 1993
Tripneustes ventricosus* 80-90 Lewis, 1958
Пор. Echinoida
Сем. Echinidae
Echinus esculentus* 35 Comely, Ansell, 1988
« 15 Gage, 1992
Loxechmus albus* 35 Bustos et al., 1991
« 20 Gebauer, Moreno, 1995
Paracentrotus lividus* 7-15 Cellano, Fenaux, 1990
« 12 Brias, LeGall (Leighton, 1995)
« 8-11 Willis (Leighton, 1995)
« 5-16 Leighton, 1995
« 8 Jangoux (персональное сообщение)
« 20 Shpigel (персональное сообщение)
Сем. Echinometridae
Echinometra mathaei 25 Drummond, 1993
Сем. Strongylocentrotidae
Strongylocentrotus 10-15 Miller, Mann, 1973
droebachiensis* 15 Fletcher et al., 1974
« 15-20 Sivertsen, Hopkins, 1995
« 5 Meidel, Scheibling, 1998
S. franciscanus * 20 Bernar, Miller 1973
« 13 Ebert, Russell 1993
S. intermedius* 15 Fuji, 1960
« <10 Kawamura, 1964
« 18 Taki, 1986
S. nudus* 30 Fuji, 1960
« 16 Kawamura, 1966
S. purpuratus* 18 Kenner, 1992
* — Виды, представляющие промысловый интерес
жизни Echinus esculentus, Strongylocentrotus purpuratus и Strongylocentrotus franciscanus, чем таковую Lytechinus anamesus, Diadema antillarum, и Tripneustes ventricosus. Результаты его исследо-
ваний (Ebert, 1982) показали, что Tripneustes gratilla, по существу, — однолетний вид, имеющий продолжительность жизни около одного года, в то время как виды типа Heterocentrotus mammillatus и Helio-cidaris erythrogramma имеют вероятность ежегодного выживания более чем 0,9. Литературные источники свидетельствуют о значительной разнице между видами в продолжительности жизни (табл. 5). Даже довольно длительные сроки продолжительности жизни, приведенные для видов сем. Strongylocentrotidae, по-видимому, являются заниженными, исходя из данных Т. Эберта (Ebert, 1988), свидетельствующих о продолжительности жизни Strongylocentrotus franciscanus в 100 и более лет. Другие авторы (Russell et al., 1998) получили близкие значения продолжительности жизни для Strongylocentrotus droebachiensis в зал. Мэн и сделали вывод, что данный вид растет медленно и является долгоживущим.
Воспроизводство
Под термином «гонадная продукция» обычно понимают репродуктивный выход (различие между максимальным и минимальным гонадным индексом), а не репродуктивные усилия (относительное количество поглощенной или адсорбированной энергии, использованной для гонадной продукции) (Lawrence, 1985, 1987). Хотя измерение количества гонадной продукции является важным моментом для понимания репродуктивного потенциала, для оценки типа стратегии необходимо определение именно относительного распределения энергии на гонадную продукцию. Таким образом, более существенным для практического использования в аквакультуре является анализ репродуктивных усилий вида.
Вид, использующий большую часть потребленной пищи на поддержание своей жизнедеятельности, будет иметь низкую эффективность ассимиляции. Хотя стресс-толерантные виды способны продуцировать крупные гонады, по эффективности они будут уступать таковым руде-ральных видов. Таким образом, хотя в литературе существуют многочисленные данные о гонадной продукции разных видов, они не могут быть применены для оценки эффективности репродуктивных усилий.
Скелетные структуры
Т. Эберт (Ebert, 1982) обнаружил, что развитие скелета (панциря и игл) и гидродинамическая экспозиция различных видов морских ежей варьируют независимо и, с другой сторо-
Таблица 4. Возраст (месяцы) и размер (диаметр панциря, мм) наступления половозрелости видов морских ежей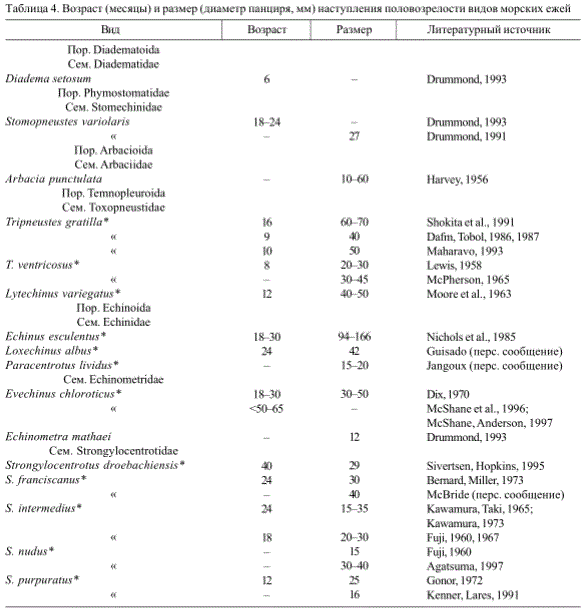
* — Виды, представляющие экономический интерес;----нет данных
ны, связаны с фактором выживания (табл. 6). При равных гидродинамических условиях в биотопах выживание было непосредственно связано с массой стенки тела. При сходных значениях масс стенки тела, выживание было непосредственно связано со степенью прибойности местообитания. А. Драммонд (Drummond, 1993) также нашел прямую корреляцию между массой стенки тела и воздействием прибоя на трех видов морских ежей. Исследования, проведенные в морях России, показали статистически достоверную корреляцию между факто-
рами среды, такими как глубина обитания и степень прибойности, и морфологическими характеристиками морских ежей р. Strongylocent-rotus — формой, толщиной и максимальным диаметром панциря, а также характером игольного покрова (Бажин, Степанов, 2002). Размещение ресурсов на структурную защиту является характерной чертой видов с низким продукционным потенциалом. Как следствие, эти виды имеют низкие темпы роста и уровень репродукции и высокие траты энергии на поддержание жизнедеятельности.
Таблица 5. Продолжительность жизни (годы) у разных видов морских ежей
Т. Эберт (Ebert, 1975), проанализировав литературные данные, сделал вывод, что вариабельность в темпах питания не связана с различиями в скорости роста и выживании видов морских ежей. Это довольно неожиданный вывод, поскольку все энергетические траты в правой стороне уравнения энергетического баланса зависят от количества потребленной энергии. Особенности, которые увеличивают способность добывать и всасывать пищу более высокого качества, по-видимому, наиболее сильно развиты у видов, реализующих рудеральную или конкурентную стратегии.
Таблица 6. Интерсепт (отрезок, отсекаемый на координатной оси) аллометричесюго отношения между сырой массой стенки тела и общей сырой массой тела видов морских ежей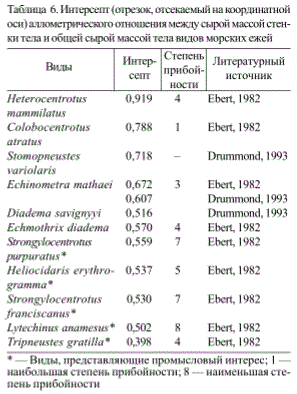
* — Виды, представляющие промысловый интерес; 1 — наибольшая степень прибойности; 8 — наименьшая степень прибойности
Способность использовать такой источник пищи как обрывки водорослей, уменьшает или даже устраняет зависимость морских ежей от продукции in situ. Tetrapygus niger, к примеру, обнаруживает наиболее высокие темпы питания при использовании инкрустирующих водорослей, в то время как L. albus наиболее эффективно питается дрифтовыми водорослями (Contreras, Castilla, 1987). L. albus имеет обилие амбулакральных ножек с большими присосками, в то время как таковые T. niger менее многочисленны и имеют минимальный размер присосок. Аристотелев фонарь Tetrapygus niger по размеру больше, чем таковой у L. albus, что непосредственно связано с пасущимся типом его пищевого поведения. На побережье Средиземноморья Paracentrotus lividus использует в пищу как прикрепленные, так и дрейфующие водоросли, другой вид — Arbacia lixula — питается инкрустирующими водорослями, а также пасется в куртинах макроводорослей (Frantzis et al., 1988). Темпы питания Lytechinus variegatus выше, чем у Arbacia punctulata (Hill,
1998). Вероятно, именно найденное различие между другими пасущимися и чисто растительноядными видами ответственно за больший выход продукции у P lividus, Tripneustes spp. и Strongylocentrotus spp.
На темпы питания оказывают воздействие температура, размер тела, физиологическое и репродук-
тивное состояние особи и даже вид пищи (Klinger, 1982). Наилучший сравнительный материал может быть получен при изучении разных видов в одинаковых условиях, как выполнено С. Контрерасом и Д. Кастиллой (Contreras, Castilla, 1987). Фенотипическая изменчивость Аристотелева фонаря — обратная зависимость между обилием пищи и его относительными размерами — была обнаружена у Paracentrotus lividus (Regis, 1978), Diadema setosum (Ebert, 1980), Echinometra mathaei (Black et al., 1982, 1984), Strongylocentrotus purpuratus (Ebert, 1982) и Diadema antillarum (Levitan, 1991). С другой стороны, подобной фенотипической изменчивости Аристотелева фонаря не было обнаружено у Strongylocentrotus franciscanus (Ebert, Russel, 1993), Tetrapygus niger (Lawrence et al., 1996) и Strongylocentrotus droebachiensis (Lawrence et al., 1998). Т. Эберт и М. Рассел (Ebert, Russel, 1993) предположили, что отсутствие вышеописанного типа морфологической изменчивости у Stron-gylocentrotus franciscanus может определяться более жесткой программой развития, чем у других видов. Другое объяснение предложил Д. Лоуренс с соавторами (Lawrence et al., 1996) для Tetrapygus niger. По их мнению, отсутствие данной изменчивости Аристотелева фонаря является характерной для стресс-толерантных видов и обусловлено более жестким контролем размеров тела.
Дыхание и обмен веществ
Анализ дыхания и обмена веществ является сложной проблемой, поскольку эти процессы вовлечены и в продукцию (анаболизм) и в поддержание жизнедеятельности (восстановление поврежденных и утраченных частей тела). Можно предположить, что у рудеральных видов преобладающая часть ресурсов расходуется на процесс анаболизма, тогда как у стресс-толерантных видов — на поддержание жизнедеятельности. Однако данные различия в путях размещения ресурсов не отражаются на общем энергетическом балансе.
Расчет связанной с дыханием энергии, так же как и темпов роста, на основе методики измерения сырой или сухой массы, может ввести в заблуждение, поскольку оба этих подхода не используют метаболически активную ткань. Использование общего количества органического вещества или общего количества белка также полностью не устраняет проблему расчета энергии, так как эти методы включают метаболически бездействующие органические соединения.
Более адекватным методом для этой цели представляется метод измерения концентрации ДНК.
Исходя из присущей каждому виду интенсивности обмена веществ, наиболее показательным может оказаться метод измерения активности метаболических ферментов.
Действительно, специфическая активность гли-колитических и гексозомонофосфат-шунтирующих ферментов была самая высокая у Lytechinus vari-egatus, промежуточная — у Echinometra lucunter, самая низкая — у Arbacia lixula (Bianconcini et al., 1980), как и можно было ожидать, исходя из других характеристик этих видов.
Высокая степень устойчивости к голоданию может рассматриваться как признак низкой интенсивности основного обмена веществ. Например, было предсказано a priori, что степень устойчивости к голоданию у особей Lytechinus variegatus будет минимальной, у Echinometra lucunter — промежуточной, и у Eucidaris tribuloides — максимальной. Особи были помещены в один аквариум с замкнутой системой циркуляции морской воды без источника пищи. Результаты эксперимента полностью подтвердили прогнозируемые различия в способности видов выдерживать голодание (рис. 1).
Вкусовые качества (съедобность)
Выметанные в процессе нереста яйцеклетки морских ежей весьма уязвимы к воздействию хищников. Одним из эффективных средств предотвращения хищничества посредством снижения съедобности особи является выработка вторичных метаболитов. Однако само по себе приобретение свойства «несъедобности» недостаточно, если это не приводит к соответствующим результатам. Р Фишер (Fisher, 1958) предположил, что если один или несколько потомков одной пары изъяты хищником из большого скопления особей, и при этом хищник приобрел 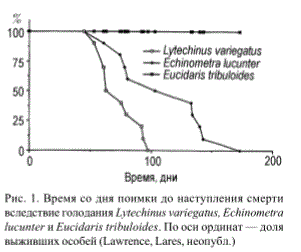
Рис. 1. Время со дня поимки до наступления смерти вследствие голодания Lytechinus variegatus, Echinometra lucunter и Eucidaris tribuloides. По оси ординат — доля выживших особей (Lawrence, Lares, неопубл.)
томков, то в процессе отбора частота гена, отвечающего за свойство несъедобности, будет увеличиваться в данном скоплении. В процессе поимки важно, чтобы хищник мог различать непригодные для питания объекты. Если принять во внимание, что ядовитые или несъедобные насекомые часто окрашены в яркие цвета, то вполне вероятно, что непригодность яиц морских ежей сем. Arbaciidae в качестве пищи обусловливается содержащимся в них фиолетовым эхинохромом полигидрооксинаф-тохиноном (McClendon, 1912). Концентрация этого эхинохрома в яйцах Arbacia punctulata достигает 0,58 г/100 мл (Ball, Cooper, 1949).
Такой тип реакции, как можно объяснить, является показательным для стресс-толерантных видов, плодовитость которых ограничена. Иными словами, подобный механизм увеличивает вероятность выживания потомства.
Пластичность в реализации стратегий
Судя по относительно большой продолжительности жизни (Ebert, 1975; Sivertsen, Hopkins, 1995; Russell et al., 1998; Ebert, 1998) и относительно низкой скорости роста (Fuji, 1960; Ebert, 1975, 1982; Sivertsen, Hopkins, 1995; и др.), стронгилоцентроти-ды в условиях наиболее типичных местообитаний, которые чаще всего характеризуются как оптимальные, реализуют первичный конкурентный тип стратегии (табл. 1). В этих условиях виды расходуют ресурсы равномерно на три базовые функции: поддержание, рост, воспроизводство. Наиболее ярким примером, демонстрирующим пластичность в реализации типов жизненных стратегий в различных условиях, является Strongylocentrotus droebachiensis. Он проявляет разнообразие адаптационных возможностей широко распространенного вида, типично обитающего в верхних горизонтах сублиторали, где факторы среды варьируют в широком диапазоне в зависимости от широтного расположения биотопа (Бажин, 1995, 2002, 2005; Бажин, Степанов, 2002). При заселении границ биотопов или границ своего ареала (где доминируют субоптимальные условия среды) этот вид подвергается стрессу абиотического характера (табл. 7). В этом случае он проявляет черты стресс-толеран-тного вида, направляя основные энергетические траты на поддержание жизнедеятельности. При этом у данного вида снижается (иногда до негативного) скорость роста (Himmelman, 1986), а продукция гонад сводится почти к нулю. Аналогичным образом вид реализует свои жизненные функции при обитании в так называемых «ежиных пустошах», где при благоприятных условиях оседания и низ-
ком прессе хищничества длительное время поддерживается высокая плотность особей (до 200-400 экз./м2). Вследствие этого наблюдается острая нехватка пищевых ресурсов и, уже как результат напряженной пищевой конкуренции, среди особей морских ежей S. droebachiensis возникает явление каннибализма.
При воздействии достаточно сильного пресса хищничества или промышленного изъятия и при отсутствии других негативных факторов происходит переключение размещения ресурсов полностью на рост и размножение. В условиях обилия пищи животные переходят к пассивному типу питания «сиди и жди». Увеличение темпов роста и гонадной продукции в этих условиях дает право обозначить данный тип стратегии как конкурентно-рудеральную.
Этот эврибионтный вид способен заселять районы с пониженной соленостью, подверженные загрязнению или эвтрофикации, где реализует вторичную конкурентно-стресс-толерантную стратегию (табл. 7).
Аналогично X droebachiensis, другие виды этого рода вполне вписываются в предложенную схему реализации функциональных особенностей при обитании в различных биотопах. Некоторая специфичность этих видов заключается в меньшем спектре условий среды и воздействующих на них экологических факторов в отличие от эвритопного S. droebachiensis.
ЗАКЛЮЧЕНИЕ
Рассмотрение видов морских ежей показывает, что они имеют набор скоординированных характеристик, которые могут быть связаны с различными типами жизненных стратегий. Эти стратегии могут быть интерпретированы на основе энергетического баланса, который свидетельствует об относительном размещении ресурсов в различные функции. Эти данные, естественно, нельзя считать заключительными, во многих случаях они просто недоступны. Нельзя ожидать, что все характеристики недвусмысленно укажут определенный тип жизненной стратегии. Не только характеристики могут взаимодействовать сложными путями, но и сами виды могут иметь вторичные жизненные стратегии, связанные с биотопами, которые отличаются по степени стресса и нарушения и приводят к проявлению промежуточных уровней этих характеристик. Необходимо учитывать, что у некоторых видов встречается фенотипическая изменчивость, которая затрудняет интерпретацию; особенно это касается конкурентных видов.
Таблица 7. Типы первичных и вторичных стратегий и связанные с ними функциональные характеристики, реализуемые морскими ежами p. Strongylocentrotus в различных местообитаниях северо-западной части Тихого океана. Модифицировано из: Lawrence, Bazhin, 1995
Условия среды Оптимальные Субоптимальные
Наличие и характер Отсутствие Биотический Биотический Абиотический стресс: Биотическое Биотическое Абиотическое
стресса или стресса и стресс и абиотический смертность молоди или нарушение: нарушение: нарушение:
нарушении нарушении стресс отсутствиеусловии для ее оседания смертность крупных особей смертность мелких особей смертность мелких особей
Обилие пищи «Ежиная Граница Пониженная соле- Высокий пресс Высокий Периодический
Характеристика и низкии пресс пустошь» биотопа, ность, загрязненный хищничества пресс мелких сброс талых или
местообитания хищничества ареала или эфтрофицирован-ный район каланов или промысла хищников загрязненных вод
Соленость + + + — + + ±
в Температура + + ± - + + ±
| ^ Гидродинамика й н Субстрат + + ± + + + +
+ + ± ± + + +
(§ Л Пища + - - + + + +
Хищничество + + + + - - +
Конкуренция + - + + + + +
Преобладающий тип Конкурентная Стресс Стресс- Конкурентно- Конку рснтно- Конкурентно- Рудерально-
стратегии (по Грайму) (С) толерантная (8) толерантная (8) стресс-толерантная (СБ) рудеральная (СИ) рудеральная (СИ) стресс-толерант- ная(Я8)
Размещение Размножение, Поддержание Поддержание Поддержание Размножение, Размножение, Размножение,
энергии поддержание жизнедеятельности, рост жизнедеятель- ности жизнедеятель- ности жизнедеятельности рост рост рост
Преобладающие размеры особей Все размерные группы или крупные Мелкие и средние Мелкие и средние Крупные Мелкие и средние Крупные Крупные
Плотность особей Высокая Очень высокая Низкая Низкая Высокая или очень высокая Низкая Низкая
Характеристика поселения Процветающее поселение Агрессивные карликовые особи Угнетенные карликовые особи Долгожители Молодые особи Долгожители Долгожители
Пищевое поведение Полупассивное Активное Активное Пассивное (сиди и жди) Пассивное (сиди и жди) Пассивное (сиди и жди) Пассивное (сиди и жди)
Гонадная продукция Высокая Низкая Низкая Низкая Высокая Высокая Очень высокая
«+» — благоприятное воздействие фактора; «-» — неблагоприятное воздействие фактора
Бажин, Лоуренс
Однако внимательное рассмотрение функциональных характеристик показывает широкое различие в первичных стратегиях. Например, токсоп-неустиды реализуют стратегию, по своим свойствам более близкую к рудеральной стратегии. Ближе к стресс-толерантной стратегии — цидаро-иды и некоторые эхинометриды, ближе к конкурентной стратегии — эхиниды и стронгилоцентроти-ды. Среди них наиболее пригодными для аквакультуры могут считаться токсопнеустиды, так как они расходуют на продукцию энергии больше, чем на защитные функции и поддержание жизнедеятельности. Как следствие этого, они быстро растут и имеют высокий уровень гонадной продукции в раннем возрасте. По сравнению с токсопнеустидами, эхиниды и стронгилоцентротиды на продукцию тратят относительно меньше энергии, в результате чего они медленнее растут и имеют меньшие репродуктивные усилия.
Высокая степень лабильности стронгилоцент-ротид в реализации разных типов жизненных стратегий, по-видимому, не является исключительным случаем среди других видов морских ежей, и, очевидно, связана с их довольно широким географическим распределением. Однако последнее, скорее всего, является не причиной, а следствием их высокой адаптационной способности. Ограничения, накладываемые со стороны относительно низкой скорости роста, не позволяют стронгилоцент-ротидам в полной мере реализовать рудеральный тип стратегии, наиболее подходящий для аквакультуры. Однако при создании соответствующих благоприятных условий, а также при искусственно регулируемой плотности, виды, по-видимому, будут способны реализовать вторичный тип стратегии, следствия осуществления которой будут вполне удовлетворять нуждам аквакультуры.
Знание основных биологических особенностей морских ежей, которые эволюционировали для максимального соответствия видов специфическим местообитаниям, является существенным как для оценки перспективности их использования в аквакультуре, так и для установления самой аквакультуры морских ежей. Необходимым условием для вида, планируемого к использованию в процессе аквакультуры, является составление его энергетического баланса. Это представляется весьма сложной и трудоемкой задачей, поскольку для этого требуется анализ, как минимум, размерных характеристик, репродуктивного цикла, температурных условий и режима питания конкретного вида. Однако эта базисная информация, которая должна применяться в аквакультуре морских ежей, отсутствует для большинства видов.
А. Г. Бажин, Д. М. Лоуренс
Зав. лаб., Камчатский научно-исследовательский институт рыбного хозяйства и океанографии 683000 Петропавловск-Камчатский, Набережная, 18
*Университет Южной Флориды, Отдел биологии, 4202 Флорида, 136 Тампа, США, 336205150 Тел., факс: 8139742549; 8139743263 E-mail:



