Общая характеристика ихтиофауны Белого моря
Составу ихтиофауны Белого моря посвящено довольно много работ. Полный список рыб с их описаниями и сведениями о распространении и биологии имеется в капитальной монографии о рыбах северных морей А. П. Андрияшева (1954). Полученные после этого новые данные позволяют лишь немного дополнить его и внести небольшие поправки.
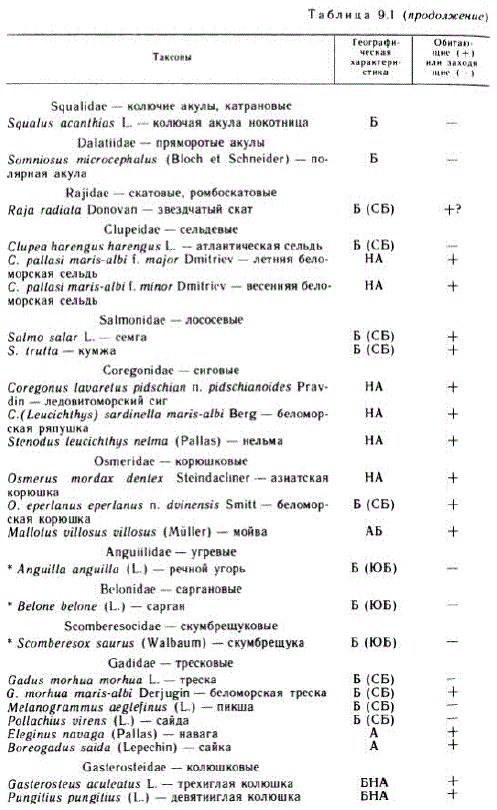
Указываемая к настоящему времени для Белого моря ихтиофауна насчитывает 56—57 видов (табл. 9.1), что почти в 3 раза меньше, чем в непосредственно сообщающемся с ним бореально-арктическом Баренцевом море (147 150 видов), и примерно столько же, сколько в арктическом Карском море (56 видов),
В Белом море не размножаются 13 видов рыб из списка (табл. 9.1). Из них 6 тепловодных, южнобореальных видов заходят в Белое море не каждый год, некоторые очень редко, в основном в годы усиления Мурманской ветви теплого Нордкапского течения. Таковы китовая акула и опах (отмечены только однажды), угорь, сарган, скумбрещука и скумбрия. Стайная рыба скумбрия заходит иногда стаями, дающими уловы промышленного значения (Андрияшев, 1954). Более или менее обычно заходят, но не размножаются, северобореальные нокотница, сайда и Пикша, морской окунь, морская камбала и молодь норвежской весенней сельди, а также нерестящаяся на глубинах открытых морей арктическая полярная акула.
Таблица 9.1
Список морских и проходных рыб и круглоротых, известных для Белого моря (по; Андрияшев, 1954; Мухомедияров, 1963а, 1963б) с некоторыми изменениями (звездочкой отмечены редко заходящие виды)


В таблице имеются некоторые изменения номенклатуры и состава ихтиофауны против имеющихся списков ихтиофауны (Андрияшев, 1954, Мухомедияров,
Как видно из табл. 9. 1, в целом наиболее богаты видами в Белом море семейства тресковых и керчаковых, представленные каждое 5—6 видами. По 4 вида содержат семейства бельдюговых и камбаловых, по 3 вида сиговые и корюшковые, лумпеновые, агоновые и липаровые. Остальные 17 семейств представлены в Белом море 1—2 видами каждое. Имеется только 7 таксономически довольно слабо, не свыше подвида, обособленных собственно беломорских или беломорско-печорских форм (Мухомедияров, 1963а): беломорская сельдь (CIиреa paІІasi тaris-albi), ледовитоморской сиг (Coregonus la varetus pildschian n. pildschiar oides), беломорская ряпушка (C. sardinella maris-albi), беломорская корюшка (Osmerus eperlanus eperlanus m. duinensis), беломopская треска (Gadиs, тогhиатаris-albi), ликод (Lиcodes pallidus maris-albi) и зубатка (Anarhichas lupus maris-albi). Столь малая таксономическая обособленность эндемиков ихтиофауны объясняется исторически малым возрастом Белого моря, которое освободилось от льдов всего за 8—12 тыс. лет до нашего времени.
Своеобразной особенностью ихтиофауны Белого моря, связанной с мелководностью входа в него и преимущественным заселением этого водоема из мелководной шельфовой зоны Ледовитого океана, является и то, что большинство семейств представлены в нем в основном шельфовыми мелководными видами. Так, семейство сиговых представлено тремя прибрежными видами, а мигрирующий несколько дальше от береговой линии омуль Coregonus autumnalis (Pallas) в собственно Белом море отсутствует, хотя он распространен вдоль всего побережья Сибири, встречается и в Мезенском заливе. Среди постоянно живущих в Белом море бореальных тресковых нет сайды и пикши, хотя есть треска, молодь которой придерживается прибрежной зоны. Беломорские виды лумпеновых, бельдюговых, керчаковых, пинагоровых, липаровых, камболовых — это в основном мелководные прибрежно-шельфовые виды распресненных солоноватых вод. Для этих семейств характерно отсутствие в Белом море обычных в Баренцевом море видов и родов более открытых вод, каковы, например, Lumpenus lampretaeformis (Wallhaum), Lycodes rossi Malmgren, Trigops nybelini Jensen, Icelus spatula Gilbert et Burke, Artediellus scaber Knipowitsch, Careproctus reinhardti Kröyer, Hippoglossoides platessoides linandoides (Bloch). Только широко распространенные в открытых водах Баренцева моря, северной части Атлантического океана и сопредельной Арктики атлантические лососи (Salто spp.), норвежская сельдь, мойва и связанная со льдами открытого моря сайка представляют собой вселившиеся в Белое море виды открытого моря.
Белое море, будучи в основном частью ареала арктических, преимущественно прибрежно-шельфовых видов, способных выдерживать пониженную соленость (Зенкевич, 1933), является в то же время краевым районом распространения бореальных видов, преимущественно атлантического генезиса, частично вселившихся, иногда временно или время от времени заходящих в него. С северо-запада беломорская ихтиофауна получила бореальные виды: звездчатого ската (Raja radiatа), атлантическую норвежскую сельдь (Clupea haren gus hareng us), атлантического лосося рода Salmo (Salmo salar, S. trutta), треску (Gadus morhus maris-albi),атлантического керчака (Muyoxocephalus Scorpius), агонуса (Agonus cataphractus), пинагора (Cyclopterus lumpus), зубатку (Anarhich as lupus maris-albi), а также временно случайно заходящие в Белое море бореальные и южнобореальные виды. С востока в Белое море вселилась преимущественно фауна шельфовых вод, представленная главным образом арктическими и нижнеарктическими видами, — миногой, нельмой и сигами, корюшкой, лумпеновыми, ликодами, агоновыми (кроме Agonus cataphractик), липаровыми, камбаловыми.
Ихтиофауны северных вод Атлантического и Тихого океанов содержат довольно много близких, амфибореальных форм (Берг. 1934: Андрияшев, 1944, 1957), разобщенных в настоящее время суровыми температурными условиями морей северной Азии и Америки, что свидетельствует о менее холодном ранее режиме этих вод: температура воды была на 2—3 °С выше 5—7 тыс. лет назад в послеледниковое литториновое потепление, и на 510 °С - в конце плиоцена (около 10 млн. лет назад) (Андрияшев, 1944. Несис, 1961). Во время ледникового периода имели место и отдельные потепления, также обеспечивало возможность распространения тихоокеанских и атлантических видов. Состав ихтиофауны Белого моря отражает эти изменения климата арктики (а также изменения водности Берингова пролива). В нее входят амфибореальные и, частично, амфиарктические компоненты разного таксономического уровня, родственные на подвидовом или видовом либо тождественные на видовом уровне (табл. 9.2), ареалы которых разобщены собственно арктическими водами Северной Америки и Северной Азии. Как видно из табл. 9.2, тождественных видов в этой группе насчитывается 6—7, представленных подвидами 5, представленных близкими видами 4. Амфибореальные виды — заметный компонент ихтиофауны Белого моря.
Таблица 9.2
Тождественные и близкие амфибореальные виды рыб, разобщенные арктическими морями Северной Азии и Северной Америки
Erelikopte x = Muppe HIL. Hij wil opg|Kir Bini i ti pi. Ti i Jiu i Eriti i R. ஃ Il 3 JIE

* Указывается для Берингова моря (Федоров, 1973). "" Указывается для Японского моря (Линдберг, Красюкова, 1987),
Таким образом, в Белом море существуют арктические виды холодных вод и бореальные умеренно холодноводные виды различного генезиса и разного исторического возраста. Это было впервые отмечено для фауны беспозвоночных (Дерюгин, 1937). Параллельное сопоставление данных о воспроизводстве и развитии близких видов и родов беломорских рыб в разных семействах представляет поэтому существенный интерес, отражая экологогеографические различия (Расс, 1977, 1986) и позволяя проследить имеющие общий характер закономерности. Такое сопоставление возможно для видов и родов ceмейств, представленных в Белом море несколькими видами (сельдевых, тресковых, керчаковых, камбаловых, а также частично пинагоровых и липаровых), с привлечением данных о близких видах из сопредельных морей. Невозможно, к сожалению, такое сопоставление для лумпеновых и бельдюговых рыб вследствие крайней недостаточности сведений об их размножении и развитии в Белом море.
Некоторые особенности строения и воспроизводства ряда видов отражены в табл. 9.3; видно, что в семействе сельдевых возможно сопоставление нескольких форм постоянно обитающей в Белом море малопозвонковой сельди (Clupea palla si maris-albi ) : Meлких, размножающихся в весеннее более холодное время при температype oт -0,8 до 0,5-4,0 °C (f. minor - no: Дмитриев, 1946) егорьевских, размножающихся летом в теплое время при 5— 10 °С онежско-сороцких сельдей, а также более крупных ивановских сельдей (f. таior). В семействе тресковых имеется возможность сопоставить холодноводные арктические виды – пелагическую сайку и природную навагу с умеренно холодноводной северобореальной треской. B полиморфном семействе керчаковых сопоставление холодноватоводных арктических видов с бореальными необходимо проводить по подсемействам, привлекая близкие виды других морей отсутствующие в Белом море, - бореального Gumnocanthus herzensteini Jordan et Starks в подсемействе Gymnocanthinae и арктического Triglops nubelini Jensen в подсемействе Tгiglopsiпае. Для сопоставления с бореальным пинагором (Cvclopteridae. Cuiclopterus /u mpu s ) привлекаются, правда с определенной натяжкой. арктические баренцевоморские виды рода Eumicrotremus (Eumicroire mus derjugins Popow, E. spinosus, Fabricius), а для сопоставления с арктическим чернобрюхим липарисом (Liparis fabricii) — более тепловодные собственно бореальный L. liparis и южнобореальный L. montegu, ошибочно считавшиеся еще недавно также обитателями Белого моря. Наконец, в семействе камбаловых сопоставляются арктическая холодноводная полярная камбала (LiopseIIa glacialis) с умеренно холодноводными речной камбалой и камбалой-ершоваткой (Platichthys esus bogdanovi, Limanda limanda).
Данные по последним шести семействам рыб Белого моря суммированы в табл. 94, рассмотрение которой показывает следующее. Отчетливо видно, что у более холодноводных видов рыб икринки крупнее, чем у родственных тепловодных. Это наблюдается как в случае более холодноводных и более тепловодных форм беломорских сельдей, так и у арктических и бореальных тресковых, керчаковых, пинагоровых, липаровых, камбаловых. При этом в семействе липаровых среди трех сопоставляемых видов самые мелкие икринки имеются у распространенного южнее двyx других видов южнобореального Liparis montagui, самые крупные — у арктического L. fabricії, промежуточные по размеру — у северобореального.


Из табл. 9.4 видно также, что более холодноводным видам, имеющим более крупные икринки, свойственно, кaк правило, большее число позвонков («правило Джордена» — см.: Расс, 1948). Однако здесь имеется несколько исключений: число позвонков, по-видимому, не различается у форм беломорских сельдей (хотя возможны различия средних), у арктических колючих пинагоров (Eитісrotгетиs sрр.) и бореального пинагора (Cyclopterus Iитриs), а у бореальной камбалы-ершоватки (Liтапda Iітапda) оно равно или немного больше, чем у арктической полярной камбалы (Liopset ta glacialis). Ecли число позвонков обычно связано с размерами икринок рыб, то размеры особей в северных водах практически не связаны с числом позвонков, Как видно из табл. 9.4, размеры особей арктических (или более холодноводных) видов сельдей, тресковых, керчаковых, пинагоровых, липаровых (?), камбаловых меньше размеров близких умеренно холодноводных бореальных видов, тогда как число позвонков у арктических видов больше (соответственно большим размерам икринок), чем у бореальных. Увеличение размеров тела у имеющих большее число позвонков (и крупнее икринки) более северных видов происходит от южнобореальных к северобореальным видам, как это показал Линдси (Lindsey, 1966, 1976), назвавший эту закономерность плеомеризмом, но у арктических видов такое явление не отмечается (Расс, 1986).
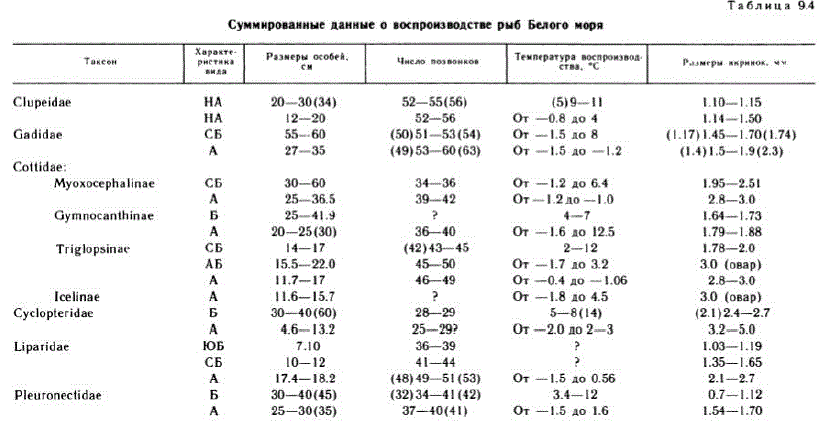
Прослеживаемое на данных о воспроизводстве рыб Белого моря увеличение размеров икринок при развитии в условиях более низкой температуры соответствует глобальному правилу
близких форм (видов, родов), установленному сначала для рыб северной части Атлантического океана (Расс, 1941), а затем подтвержденному данными по разным районам Мирового океана (Андрияшев, 1954, Перцова-Остроумова, 1961; Пермитин, Сильянова, 1971; Расс, 1977, 1986). Указанное «правило размеров яиц» существует, по-видимому, для всех холоднокровных животных — рыб, водных беспозвоночных (ракообразных, моллюсков), амфибий и рептилий Северного полушария (Расс, 1985) при малом числе исключений.
Как видно из изложенного, исследованная ихтиофауна Белого моря представляет большой интерес для познания истории этого водоема, истории населяющей его фауны, эколого-географических взаимоотношений ее компонентов. Особенно перспективными представляются сравнительные исследования воспроизводства и развития рыб и беспозвоночных Белого моря, совершенно недостаточно изученные до настоящего времени.



