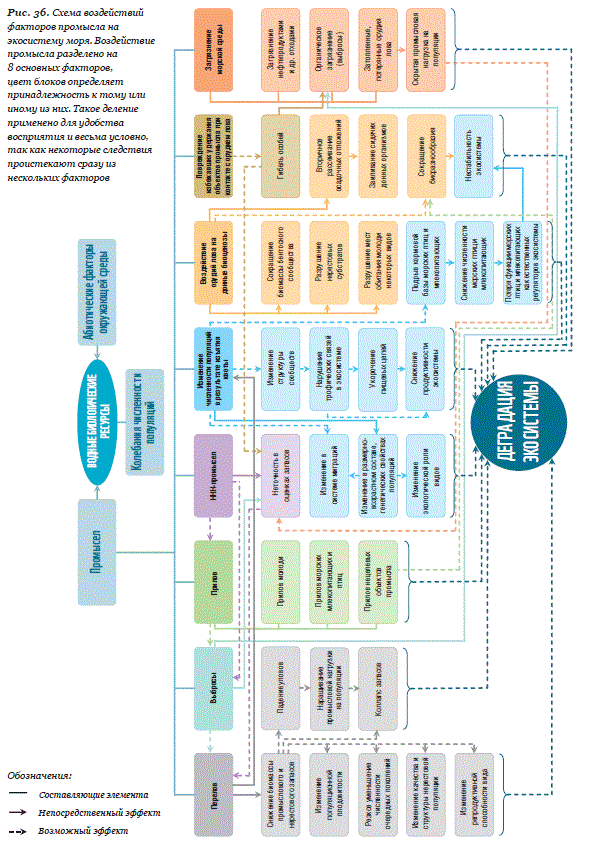
Промысловое изъятие в рамках научно обоснованного ОДУ
Самым очевидным и, возможно, наиболее существенным воздействием
рыболовства на коммерчески значимые виды рыб на сегодняшний день
является предусмотренная эксплуатация запасов, которая сопровождает-
ся изменением численности популяций.
Крупномасштабное промысловое изъятие любого вида из экосистемы,
которая эволюционировала в течение многих тысячелетий, не может
происходить бесследно для этой экосистемы. С одной стороны, уже
очевидно, что последовательное применение парадигмы максимально-
го устойчивого вылова (MSY) не гарантирует сохранение экосистем, с
другой – настолько же очевидно, что невозможно полностью отказать-
ся от использования морских биологических ресурсов. Вопрос только
в том, насколько мы можем эти изменения предсказывать и насколько
они приемлемы для нас при тех объемах изъятия, которые необходимы
человеку. В настоящее время можно говорить о следующих изменениях в
экосистемах, связанных с легальным промысловым изъятием.
Генетическая структура популяций. Снижение численности вызывает
изменения в размерном и возрастном составе, генетических свойствах
и смертности популяций. Рыболовство может влиять на генетическое
разнообразие популяций и на эволюционные процессы всех видов рыб,
в том числе и непромысловых, так как модифицируются отношения
между ними и размерный состав самих видов. Жесткая промысловая
нагрузка и избирательная добыча самых крупных особей популяции
может привести к увеличению доли быстрорастущих и созревающих при
меньших размерах особей. Тем не менее, в настоящее время не имеется
достаточной информации о том, насколько значительны эти послед-
ствия для генетической структуры популяций и какое влияние они могут
оказывать на способность приспосабливаться к изменениям условий
окружающей среды (Доклад …, 2006).
Морские птицы. Промышленное рыболовство
оказывает значительное влияние на различные
компоненты экосистемы. Хотя прямое воздей-
ствие промысла на морских птиц и существует
– в частности, рыбодобывающий флот в Барен-
цевом море пугает птиц в период гнездования
и негативно сказывается на их репродукции,
основное воздействие рыболовства является
опосредованным через изменение кормовой
базы. От численности сельди, мойвы, песчанки
зависит успех зимовки и весеннего выкарм-
ливания птенцов наиболее массовых морских
птиц – чаек-моевок и кайр. Популяция тупика сильно зависит от доступ-
ности мелких пелагических рыб, особенно до и во время гнездования, в
связи с чем считается, что промысел является основной опосредованной
угрозой для популяций тупика (Предложения, 2005). На востоке Ба-
ренцева моря для многомиллионных колоний морских птиц основным
кормом является сайка. При естественных колебаниях численности
сельди, мойвы, песчанки и сайки снижение или отсутствие одного из
этих видов компенсировалось другими. Следствием резкого сокращения
запасов мойвы стало уменьшение численности молодого поколения и
популяции в целом у моевок и других чайковых птиц: особенно сильно
это отразилось на кайрах Мурманского побережья. В отдельные годы пе-
риод размножения у крупных чаек и моевок смещался на более поздние,
по сравнению со средними, сроки. У моевок и бакланов средний раз-
мер кладки находится в непосредственной зависимости от состояния их
кормовой базы: в случае субоптимальных кормовых ресурсов ее размеры
заметно ниже. У других птиц отсутствие или недостаточность необходи-
мых кормовых ресурсов в период выращивания птенцов приводит к их
массовой гибели от истощения (Краснов, 1989, 1991; Краснов и др., 1996).
Морские млекопитающие. Катастрофическое снижение численности
мойвы, сайки, сельди и других пелагических рыб существенным образом
усугубило уже развивавшиеся ранее негативные явления в состоянии
популяций морских млекопитающих. Резкое сокращение численности
рыбы принципиально изменило миграционные пути и особенности оби-
тания многих тюленей Баренцева моря (Попов, 1989). Лишившись сайки
– основного корма в период миграции, гренландские тюлени беломор-
ской популяции были вынуждены сменить пути миграций и с середины
1970-х гг. в период нереста мойвы стали появляться у северных берегов
Кольского полуострова. Однако уже в 1986–1987 гг. резкое ухудшение
кормовой обстановки привело к прекращению и этих кормовых мигра-
ций. Тюлени стали оставаться вблизи берегов Мурмана на все лето. С на-
чала зимы около полумиллиона гренландских тюленей ушли к фьордам
Норвегии, где тысячи особей погибли от истощения (Матишов,1992).
Миграционное поведение. Под воздействием промысла в 1950–1970-е гг.
изменение структуры популяции трески (резкое сокращение числен-
ности старших возрастных групп) вызвало уменьшение протяженности
миграционных путей и сокращение нагульного ареала рыб. В результа-
те треска перестала посещать восточные районы моря, где ранее особи
старших возрастов откармливались сайкой и ее молодью, люмпенуса-
ми и крабами, а пикша перестала заходить в основные районы своего
прежнего откорма в юго-восточной части моря. Все это привело к утрате
промыслового значения тех районов, которые посещались в процессе
нагульных миграций рыбами старших возрастов, и недоиспользованию
популяциями кормовой базы продуктивных мелководий на востоке Ба-
ренцева моря (Маслов, 1968; Ярагина и др., 1996б).
Рост вылова атлантическо-скандинавской сельди почти в 2,5 раза, наря-
ду с появлением неурожайных поколений, привел к столь масштабному
уменьшению нерестового запаса, что с начала 1970-х гг. сельдь перестала
совершать нагульную миграцию в открытую часть Норвежского моря,
выходить на зимовку к восточным берегам Исландии. Эта пелагическая
рыба в течение всего года оставалась в узкой (12-мильной) зоне при-
брежных вод Норвегии (Матишов, 1992).
Размерно-возрастная структура популяций. Вследствие своей селек-
тивности промысел, изымая часть популяции, меняет ее размерно-
возрастной состав. Характер этого воздействия хорошо прослеживается на
многолетнем материале по уловам трески. Преобладающими возрастны-
ми группами в уловах 1930-х гг. были 6–8-годовики, в 1950-е – 4–6-годо-
вики, причем доля самых старших рыб (от 9 лет и старше) уменьшилась
с 10,3% в 1930-е гг. до 3,5% в 1950-е (Маслов, 1968). По мере дальнейшего
развития и увеличения интенсивности промысла удельный вес старших
возрастных классов продолжал снижаться, а 3–5-годовиков – возрастал.
В 1960-е гг. уже до 70% уловов приходилось на рыб в возрасте 3–5 лет,
в 1970-е – 75% (Бенко, Пономаренко, 1972). Кроме того, к 1970 г. практиче-
ски исчезли рыбы старшего (15–20 лет) поколения, к 1980 г. почти не
стало 10–15-летней трески. Именно потеря рыб старших возрастных групп
стала самым явным негативным следствием селективного рыболовства
в Баренцевом море. Эти рыбы имеют не только высокую индивидуальную
плодовитость, но и продуцируют более крупную икру с большим запасом
питательных веществ в желточном мешке и, следовательно, с более
жизнестойкими личинками (Матишов, 1992). Возрастная структура
популяции несколько изменилась с установлением лимитов вылова
в 1980-е гг. (особенно с ограничением промысла щадящим уровнем ОДУ
в 1989–1991 гг.), после чего в стаде произошло увеличение доли особей
старших возрастных групп (Ярагина и др., 1996а).
Репродуктивная структура популяций. Интенсивный промысел ока-
зал заметное селективное влияние на половую и возрастную структуру
популяций трески. Если в 1930–50-е гг. более половины особей трески
становились половозрелыми в 10–11 лет, то в 1970-е – в 8–9 лет (По-
номаренко и др., 1985). К началу 1980-х гг. резко увеличились темпы
полового созревания, и уже к началу 1990-х гг. отмечалось до 5–20% по-
ловозрелой трески в возрасте 3, 4 и 5 лет (Матишов, 1992).
Темпы полового созревания окуня-клювача за период 1966–1982 гг. так-
же увеличились. Если в начале периода 70% созревших самок имели воз-
раст 14 лет, то в конце периода такое же их количество было в возрасте
12 лет (Матишов, 1992). К тому же под воздействием промысла особенно
уменьшилась продолжительность жизни именно самок окуня-клювача.
Если в 1950–60-е гг. в промысловом стаде преобладали самки длиной в
среднем 38 см и возраста 18 лет, то уже в начале 1980-х гг. наибольшую
долю имели особи в среднем 11-летнего возраста и длиной 30 см (Соро-
кин, Шестова, 1988).
Популяционная плодовитость. Воспроизводительную способность рыб-
ных стад, как известно, отражает видовая, индивидуальная и, в большей
степени, популяционная плодовитость. Популяционная плодовитость
трески резко сократилась за период с конца 1940-х до начала 1980-х гг. в
связи со снижением численности старших возрастных групп, индивиду-
альная плодовитость которых максимальна (Серебряков и др., 1984). Что
касается сельди и мойвы, имеющих короткий жизненный цикл, то для
них исследователи отмечали уровень популяционной плодовитости зна-
чительно ниже критической отметки на протяжении ряда лет с середины
1960-х и начала 1980-х гг. соответственно (Норвилло, 1991).
Трофические связи. Нарушение трофических связей в экосистеме, как уже
отмечалось, может иметь следствием укорочение пищевых цепей. В 1980-х
гг. баренцевоморская экосистема относительно нормально функциони-
ровала до уровня фитопланктона, а далее через пелагические сообще-
ства энергия вторичной продукции почти не утилизировалась, поскольку
численность основных потребителей зоопланктона (сельди, мойвы, сайки,
креветки) существенно сократилась. Пищевая цепь «зоопланктон – хищ-
ные донные рыбы» была несравненно короче, чем цепь, которая включала
пелагических рыб. Эвфаузииды, а также калянус, являвшиеся кормом этих
рыб, в достаточной мере не утилизировались. Объемы неиспользованной
пищи, по-видимому, могли достигать 50–60 млн т (Матишов, 1992). Таким
образом, при сложившейся на то время ситуации значительное количество
продукции низких трофических уровней не могло быть использовано про-
мыслом, поступая непосредственно в детритную цепь.



