Органические вещества
Органические вещества (ОВ), содержащиеся в воде, крайне важны для жизнедеятельности гидробионтов. Они играют значимую роль не только в процессах питания водных организмов, но и в их коммуникации (Хайлов, 1971;
Кулаковский, 2000 и др.). Кроме того, по их концентрации можно судить об интенсивности и соотношении продукционно-деструкционных процессов в водоеме. Концентрация и состав ОВ зависят также и от их поступления и выноса при водообмене. В связи с тем, каков источник происхождения органики, принято выделять автохтонный (местный) и аллохтонный (приносной) компоненты. Они обычно различаются по составу.
ОВ, содержащиеся в морской воде, представляют собой сложную смесь соединений, состоящую не только из продуктов жизнедеятельности гидробионтов, но и из преобразованных нефтяных и других соединений. Последние чаще поступают в водоем из антропогенных источников, а также при эрозии донных осадков.
Химический состав органических веществ, обнаруживаемых в воде, весьма разнообразен. Основная доля приходится на углеводы, аминокислоты и липиды. В зонах воздействия стока речных вод возрастает количество взвешенной органики и повышается содержание гуминовых кислот и фульвокислот.
Исходя из размеров, все органические вещества разделяют условно на две группы: взвешенные (ВОВ) – более 0.5 мкм и растворенные (РОВ) – менее 0.5 мкм. Их соотношение в морской воде, равное обычно 1:10, может существенно изменяться как в одну, так и в другую сторону (Виноградов, 1967). Оба эти компонента органики не только по-разному представлены в море, но и имеют разное значение для обитающих в нем организмов.
Длительное время считалось, что органические вещества, содержащиеся в воде, являются лишь объектом бактериальной переработки, в результате которой происходит пополнение запаса биогенных элементов, а прямого вклада в баланс энергии в водных экосистемах они не дают. В течение нескольких последних десятилетий показано, что такой подход не соответствует действительности. Оказалось, что РОВ поглощают и утилизируют не только микроорганизмы, служащие, в свою очередь, источником пищи для микрозоопланктона и некоторых других организмов. Из пула растворенных органических веществ напрямую черпают энергию погонофоры, кишечнополостные, полихеты, моллюски, иглокожие и многие другие морские беспозво
ночные. Особенно важен этот источник пищи для сидячих (седентарных) животных и представителей интерстициальной фауны. Многие микроводоросли также могут переходить с автотрофного на гетеротрофный тип питания, сорбируя РОВ (подробнее этот вопрос будет обсужден ниже, в главе о планктоне).
Новое видение морскими биологами места РОВ в балансе вещества и энергии в водоемах четко сформулировал В.Д.Федоров (1987), сделавший вывод о том, что органика «...играет роль резервуара субстратов и источников энергии, доступных всем осмотрофным организмам».
Полностью разделяя эту точку зрения на место растворенной органики в балансе вещества и энергии в водных экосистемах, не могу не отметить, однако, что термин «осмотрофные организмы» нельзя использовать для обозначения рассматриваемого явления, хотя это делается многими исследователями (см., например, Pandian, 1975), и ситуацию, по-видимому, уже не исправить.
Почему я на этом настаиваю? Дело в том, что об осмотическом питании можно говорить лишь в том случае, когда имеет место пассивный перенос через клеточную мембрану по градиенту концентрации. В рассматриваемом случае растворенные органические вещества перемещаются внутрь организма против огромного концентрационного градиента, что возможно лишь путем активного мембранного транспорта либо с помощью пиноцитоза (т.е. транспорта веществ в мембранной упаковке). Кроме того, в случае осмоса перемещается растворитель, т.е. вода, а не растворенные в ней вещества, которые должны задерживаться клеточной мембраной.
Большинство морских организмов – осмоконформеры. Осмотическое давление их внутренней среды равно таковому во внешней среде (морской воде). У этих животных нет эндосмоса воды, и, следовательно, говорить об их «осмотическом питании» нельзя.
В связи с обсуждением роли растворенной органики в водоемах будет нелишним вспомнить о старом споре между А. Пюттером и А. Крогом по поводу «осмотического питания» и его роли в энергетике организмов. Затронутая ими проблема широко обсуждалась и вошла во многие руководства и учебники (см., например, Зернов, 1934).
А. Пюттер (Putter, 1909) был первым исследователем, почти 100 лет назад утверждавшим, что РОВ могут абсорбироваться поверхностью тела водных организмов и служить для них источником энергии. А. Крог (Krogh, 1931) подверг эту идею жесткой критике, в основном из-за методических ошибок А. Пюттера. Известность и авторитет А. Крога сыграли негативную роль в развитии дальнейших исследований этого направления. В результате идея поступления растворенной органики в организм гидробионтов не через пищеварительную систему, а через поверхность тела, верная по своей сути, была надолго похоронена.
Однако время показало, что А. Пюттер был прав. Основную роль в реанимации его представлений сыграли результаты многочисленных исследований, начавшихся в 60– 70-х годах прошлого столетия. Среди них на первом месте стоят работы Г. Стефенса c соавторами (Stephens, Schinske, 1961; Stephens, Vircar, 1966; Stephens, 1968, 1975 и др.). Объектами этих исследований были представители 35 родов различных морских беспозвоночных: кишечнополостных, губок, моллюсков, ракообразных, полихет и иглокожих. Не детализируя полученные данные, выделим (Pandian, 1975) основные результаты работ этого направления:
- Поступление РОВ в организм морских беспозвоночных происходит через поверхность тела и ряда органов, например ктенидиев моллюсков. Поступление растворенной органики через желудочно-кишечный тракт сравнительно невелико. У различных ракообразных интенсивность поглощения растворенных веществ поверхностью тела и жабрами значительно ниже таковой у остальных исследованных беспозвоночных животных. 2. Основу абсорбции свободных аминокислот, полисахаридов и других растворенных органических веществ составляет их активный транспорт, который представляет собой энергетически зависимый процесс, блокируемый 2,4-динитрофенолом. Транспорт РОВ осуществляется против градиента концентраций, разница которых во внутренней и наружной средах достигает нескольких порядков. На него расходуется от 3 до 15% энергии, получаемой морскими беспозвоночными от утилизации абсорбированных органических соединений.
- Сорбированные органические вещества используются на синтетические процессы и покрытие энергетических трат организма. На примере некоторых актиний (Fungia scutaria и Anemonia sulcata) показано, что поглощение глюкозы из внешней среды покрывало расходы энергии на стандартный обмен, а сорбция глицина давала около 40% от общей метаболической энергии организма. В первом приближении рассчитано, что гетеротрофная утилизация РОВ обеспечивает морским беспозвоночным количество энергии, эквивалентное 50% того, что продуцируется фитопланктоном. 4. Снижение солености внешней среды приводит к угнетению абсорции РОВ.
На этом последнем пункте стоит остановиться и обсудить его подробнее. Сам по себе феномен угнетения абсорбции растворенной органики различными морскими организмами при снижении солености среды был зарегистрирован многими исследователями (Stephens, Vircar, 1966; Shick, 1973; Anderson, 1975 и др.). Получены такие данные и в экспериментах на беломорских моллюсках Mytilus edulis (Хлебович и др., 1982).
Механизмы активного транспорта растворенных органических веществ были наиболее детально исследованы на примере поступления свободных аминокислот в клетки морских моллюсков Сеrastoderma edule, Mya arenaria, Modiolus demissus и Mytilus californianus. Показано, что абсорбция аминокислот клетками жабр этих беспозвоночных подавляется по мере снижения как солености морской воды, так и содержания ионов натрия во внешней среде (рис. 7). Кроме того, поступлению РОВ из воды в клетки противостоит их отток обратно во внешнюю среду, который в норме значительно меньше абсорбции органических веществ (Ferguson, 1971). По мере снижения солености (осмотического давления) выход свободных аминокислот из клеток моллюсков резко усиливается. Показано (Pierce, Greenberg, 1973; Bartberger, Pierce, 1976 и др.), что регуляция выхода аминокислот из клеток моллюсков осуществляется
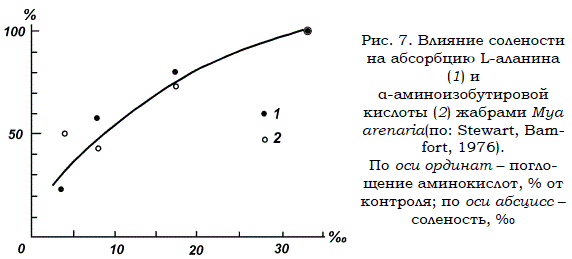
Рис. 7. Влияние солености на абсорбцию L-аланина (1) и α-аминоизобутировой кислоты (2) жабрами Mya arenaria(по: Stewart, Bamfort, 1976). По оси ординат – поглощение аминокислот, % от контроля; по оси абсцисс – соленость, ‰
специфическими мембранными сайтами, находящимися в двух конформационных состояниях: открытом и закрытом. При снижении солености возрастает доля открытых сайтов, что приводит к усиленной потере аминокислот. Таким образом, при низкой солености исследованные организмы, во-первых, не могут абсорбировать растворенные органические соединения с достаточно большой скоростью и, вовторых, усиленно теряют их из-за соответствующих изменений проницаемости клеточных мембран.
Отсюда следует также вывод о том, что абсорция РОВ как источник вещества и энергии оказывается невозможной в пресных водах. Этим обстоятельством, в частности, можно объяснить и тот факт, что поступление растворенных органических веществ через покровы тела гидробионтов не учитывается гидробиологами при анализе процессов обмена вещества и энергии, происходящих в пресноводных экосистемах.
Взвешенные частицы делятся на две составляющие: неорганические и органические вещества (Хорн, 1972). Состав органической компоненты весьма сложен. Анализировать его также сложно, поскольку нет ясности, а что, собственно, считать взвешенной органикой. Теоретически сюда попадают не только кусочки водорослей или иных отмирающих и разрушающихся организмов, но и фекалии и псевдофекалии различных беспозвоночных, образующие вместе детрит, а также и многие другие компоненты взвеси, прежде всего мельчайшие планктонные организмы. Вместе с детритом они образуют сестон, состав которого, строго говоря, не определен. Трудно ответить на вопрос о том, где та минимальная размерная граница, выше которой организмы не считаются частью сестона. Так или иначе, принято считать (Rilley et al., 1964), что на долю живых планктонных организмов приходится около 10%, а остальная часть взвеси состоит в основном из детрита, значительную долю которого составляют коллоидные органические материалы. Взвешенная органика является существенным компонентом питания различных морских беспозвоночных, как ведущих сидячий образ жизни, так и подвижных. В процессе питания различные сестонофаги тем или иным способом улавливают взвешенные в воде либо недавно осевшие на дно частицы органики, а затем перемещают их к специальным органам, где пища отделяется от воды, концентрируется и поступает в пищеварительную систему (Pandian, 1975).
Пути поступления органических веществ в водоем, как отмечалось выше, различны. В процессе жизнедеятельности различных водорослей и животных в воду выделяется огромное количество разнообразных метаболитов, создающих пул автохтонных ОВ.
К.М. Хайлов (1964а,б) высказал предположение о том, что основное количество растворенного в прибрежной морской воде органического вещества образуется за счет прижизненного экскретирования макрофитами. Эту точку зрения, по крайней мере, частично подтверждают данные о том, что Laminaria digitata и Fucus vesiculosus выделяют в морскую воду 30–35% от количества синтезированных в процессе фотосинтеза органических веществ (Sieburth, 1969). Конечно, эти факты нельзя считать доказательством того, что макрофиты выделяют большую часть растворенной органики, содержащейся в морской воде, но то, что они участвуют в этом процессе и при том достаточно значимо, можно считать установленным.
Десятью годами позже аналогичные факты были получены в экспериментах на большом числе тихоокеанских видов сублиторальных и литоральных макрофитов (Пешеходько, Титлянов, 1979). Исследовалось выделение растворенной органики такими водорослями, как Ulva fenestrate и Codium fragilie (Chlorophyta), Rhodymenia stenogona, Rhodomela larix, Ahnfeltia tobuchiensis и Grateloupia turuturu (Rhodophyta), Punctaria plantaginea, Laminaria japonica и Sargassum pallidum (Phaeophyta). Было показано, что изученные водоросли выделяют во внешнюю среду от 2 до 20% от количества углерода, ассимилированного в процессе фотосинтеза. При этом оказалось, что на свету экскреция органических соединений происходит в 2.5–3 раза интенсивнее, чем в темноте.
Сходной экскреторной активностью обладают также планктонные морские одноклеточные водоросли. В процессе роста они выделяют во внешнюю среду от 3 до 25% ассимилированного углерода (Hellebust, 1965; Samnel et al., 1969). Этот источник поступления органики за счет жизнедеятельности водорослей Белого моря не исследован, но очевидно, что описанное явление универсально. Вопрос может заключаться лишь в том, больше или меньше метаболитов выделяют беломорские макрофиты по сравнению с теми же видами из Баренцева моря и других морей Мирового океана, в отношении которых такие данные получены (Хайлов,1964 а,б; Sieburth, 1969).
В противоположность водорослям беломорские беспозвоночные, пусть недостаточно, но были исследованы с этой точки зрения. Рассмотрим в качестве примера результаты работ, выполненных Э.Е. Кулаковским, В.Н. Галкиной, Ю.И. Ляхиным и другими исследователями для того, чтобы выяснить, как поселения беломорских мидий воздействуют на окружающую среду. При этом они собирали материалы в местах как естественных, так и искусственных поселений этих двустворчатых моллюсков, культивируемых в Кандалакшском заливе (Кулаковский, 2000).
Полученные данные (табл. 4) показывают, что мидии выделяют во внешнюю среду количество растворенных органических веществ, пропорциональное скорости метаболизма, о которой судили по потреблению кислорода. При этом скорость выделения РОВ, отнесенная к весу моллюсков, была максимальной у мелких мидий (возраст 1+) из искусственных поселений.
В результате жизнедеятельности моллюсков в таких поселениях концентрация растворенных органических веществ увеличивается в несколько раз по сравнению с таковой на участках акватории, где не было мидиевых плантаций (табл. 5).
Таблица 4 Сравнение показателей жизнедеятельности мидий из искусственных и естественных поселений (с сокращениями по: Кулаковский, 2000)
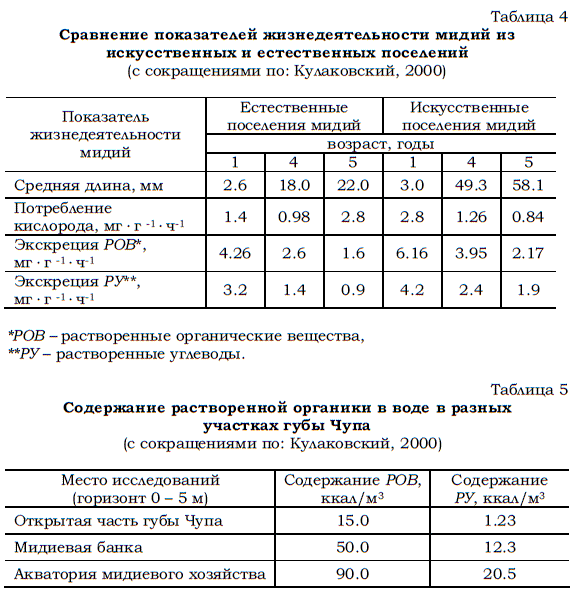
*РОВ – растворенные органические вещества, **РУ – растворенные углеводы.
Таблица 5 Содержание растворенной органики в воде в разных участках губы Чупа (с сокращениями по: Кулаковский, 2000)
Пересчитанные (исходя из содержания углерода) в соответствующие величины калорийности эти данные дают возможность оценить энергетический вклад, который осуществляют мидиевые поселения, продуцируя РОВ. Естественно, что для того, чтобы судить, сколько растворенной органики поступает в воды всего моря и каков ее энергетический вклад, этих данных недостаточно, однако они позволяют хотя бы приблизительно представить масштаб рассматриваемого явления.
Кроме того, проведенные наблюдения показали, что в районе мидиевых плантаций как на Баренцевом, так и на Белом море значительно возрастает обилие фитопланктона, а его продукция увеличивается в несколько (до 7) раз (Головкин и др., 1979). Причиной этого служат различные органические соединения и биогенные элементы, выделяемые мидиями во внешнюю среду. Подсчитано, что за час в пересчете на 1г сухой массы мидий в воду выделяется в среднем до 222 мкг органического и 13.7 мкг аммиачного азота, 40 мкг органического и 27 мкг минерального фосфора. Авторы этой статьи считают, что зоны моря вблизи мидиевых поселений (как естественных, так и искусственных) могут рассматриваться «...как триггерный механизм биогидрохимической системы прибрежья, способствующий возникновению новых вспышек развития фитопланктона в условиях низкого содержания биогенных элементов» (Головкин и др., 1979, с. 76).
Мидиевые поселения производят также и большое количество взвешенной органики. Приведем некоторые конкретные данные. Специальными исследованиями, выполненными в Кандалакшском заливе в местах культивирования мидий (мыс Картеш, Сон-остров, Иванов-наволок), показали (Кучаева и др., 1988), что взвесь состояла из фито-, зоопланктона и детрита. Фитопланктон был представлен диатомовыми водорослями. В зоопланктоне преобладали веслоногие рачки, личинки различных животных, а также простейшие. Детрит был представлен бесформенными органическими остатками и фекалиями мидий. Содержание белков в исследованной взвеси варьировало от 13 до 46% от сухой массы. Концентрации углеводов и жирорастворимых веществ составляли соответственно 2–30 и 6–28%.
Данные, полученные М.В. Ивановым (2006) в районе нескольких мидийных плантации, находящихся в Кандалакшском заливе, показали, что в среднем культивируемые моллюски выделяют около 60 кг сухих органических веществ с 1га плантации в сутки. За год такое хозяйство с максимальными биомассами мидий на субстратах создает нагрузку на окружающие экосистемы в 6–7 т сухих органических веществ. Следует при этом отметить, что хотя эти показатели достаточно велики, но они значительно меньше аналогичных данных, полученных в других морях. Об этом свидетельствуют результаты измерения скорости оседания взвешенных органических веществ, продуцируемых мидийными хозяйствами (табл. 6).
Таблица 6 Осадконакопление под плантациями по культивированию моллюсков (с сокращениями по: Иванов, 2006)

В составе ВОВ, производимых мидиями, как уже отмечалось выше, содержится большое количество фекалий и псевдофекалий. Эти продукты жизнедеятельности (не только мидий, но и других гидробионтов) служат не менее важным источником органических веществ.
Лучше всего эта составляющая баланса органических веществ в море исследована на примере планктонных ракообразных. Показано, что их фекальные пеллеты играют существенную роль в вертикальном переносе органического и неорганического вещества в водных экосистемах. Так, в некоторых районах Мирового океана до 92% органического углерода во взвешенном веществе приходится на пеллеты (Graf, 1989). Они формируют часть донного биогенного осадочного материала (Корнеева и др., 1992; Романкевич, Ветров, 2001). Пеллеты могут служить важным источником пищи для бентосных и бенто-пелагических животных, так как содержат значительное количество органических веществ, защищенных от размывания перитрофической мембраной (Atkinson, Wacasey, 1987; Urban-Rich, 1997; Mayzaud et al., 1998).
В Белом море в сестоне (размерная фракция от 1 до 76 мкм) с конца мая по середину сентября в среднем содержалось 206 ± 82 мкг органического углерода в 1 л воды, а сухой вес сестона варьировал от 0.64 до 3.65 мг/л (Мартынова, 2004). В энергетическом выражении это количество сестона соответствует приблизительно 2 кал/л. Исходя из этих данных, можно подсчитать, что сестон всего моря в этот период содержал около 1.1.106 т органического углерода, что эквивалентно 1.1.1013 ккал.
Та часть использованного в пищу сестона, которая не утилизирована планктонными ракообразными и удалена из их организма в воду в виде фекальных пеллет, имеет весьма значительную биомассу. Данные о содержании пеллет в водной толще, а также скорости их оседания были получены с помощью седиментационных ловушек, которые устанавливались в Кандалакшском, Онежском и Двинском заливах (Мартынова, 2004). В разные сезоны безледного периода в Белом море сухой вес фекальных пеллет, образованных под единицей площади морской поверхности, колеблется от 283 мг/м2 в сутки (май, глубоководные районы Кандалакшского залива) до 0.63 мг/м2 в сутки (октябрь), постепенно уменьшаясь в течение этого времени. Максимум пеллетного потока приходится на конец мая–начало июня, когда крупные копеподы Calanus glacialis активно выедают фитопланктон в период его массового весеннего цветения. В течение всего поздневесеннего и летнего периода (июль–август) пеллетный поток во всех заливах примерно одинаков: сухой вес – 37.75 мг/м2 в сутки. При этом большая часть пеллет не достигает дна и некоторое время, вплоть до своего полного разрушения, удерживается в слое скачка плотности (пикноклина). Из того количества пеллет, которые произведены планктонными копеподами в поверхностном 10-метровом слое, в неразрушенном виде дна достигает не более 1%.
Значительная часть органических веществ поступает в море с речным стоком. Это – так называемая аллохтонная органика. Основная часть аллохтонных органических веществ (порядка 60%) поступает в Белое море в период весеннего паводка, около 30% – в летне-осенний период, а оставшиеся 10% приходятся на зиму (Максимова, Владимирский, 1982). В целом за год в бассейн Белого моря поступает около 9.4 млн. т аллохтонных ОВ. С водами Баренцева моря приносится около 4 млн. т и примерно столько же выносится обратно. С речным стоком поступает около 5.4 млн. т (Максимова, 1991).
Воды Баренцева моря, поступающие, как отмечалось ранее, в основном в зимний период, опускаются на глубину и заполняют котловину моря, снабжая обитателей арктической водной массы органикой, биогенными элементами и кислородом. Поскольку обмен между морями сбалансиро
ван, а поступление органики скомпенсировано оттоком, то ее общая концентрация в водах Белого моря меняется не очень значительно.
В эстуариях при смешивании речной и морской воды за счет деятельности так называемых маргинальных фильтров (Лисицин, 1994) из оборота выводится основное количество взвешенных форм различных веществ, в том числе и органических, поступающих в море с речным стоком. Лавинное осаждение речной взвеси в эстуариях рек происходит в три основных этапа.
1 этап Подпруживание устья морскими водами и снижение скорости речного потока приводит к механическому осаждению взвеси. Этот механизм начинает действовать уже в нижнем течении рек до их впадения в море. Здесь обычно осаждаются песчаные и крупноалевритовые фракции взвеси, образуя в дельте реки отмели и острова.
2 этап В осаждении оставшихся пелитовых и мелкоалевритовых фракций (в значительной степени – коллоидных) и растворенных органических веществ основную роль играет электролитическое воздействие морской воды. В пределах солености от 5 до 15‰ происходит перезарядка коллоидов, при которой частицы проходят изоэлектрическую точку и теряют заряд. Это приводит к флоккуляции коллоидов – образованию хлопьев, получивших название «эстуарного снега». Его выпадение обусловлено не гравитационными, как на первом этапе, а коллоидными, физико-химическими законами. Именно в этой части маргинального фильтра, на этом этапе его деятельности выпадает значительная часть растворенной органики и железа.
Образующиеся хлопья содержат также частицы минеральных коллоидов – глинистых минералов, склеенные массой из новообразованных оксигидратов железа и органики. Эти флоккулы сорбируют из воды биогенные элементы, тяжелые металлы и многие загрязняющие вещества. Главными сорбентами при этом служат глинистые минералы, органические вещества (гуминовые кислоты и фульвокислоты) и оксигидраты железа.
Флоккулы становятся объектом бактериальной переработки (Мицкевич, Намсараев, 1994) и источником пищи для зоопланктонных фильтраторов. В желудках планктонных копепод, пойманных осенью в эстуариях рек Оби и Енисея, содержался только «эстуарный снег» (Виноградов и др., 1994).
В зонах эстуарных фронтов, где при солености выше «критической» (Хлебович, 1971), т.е. между 5–8 и 15‰, происходит наиболее интенсивное осаждение органических веществ, скапливается также и множество детритофагов, как бентосных, так и планктонных (Виноградов и др., 2000).
3 этап После прохождения первых двух этапов содержание взвеси в воде резко снижается, а прозрачность повышается. Как следствие этого значительно увеличиваются обилие и фотосинтетическая активность фитопланктона, а затем и обилие зоопланктона (Ведерников и др., 1994). Эта часть маргинального фильтра – биологическая. В ней образуется новая «взвесь» из той части растворенной органики, которая преодолела предыдущую сорбционную часть фильтра. Зоопланктонные фильтраторы располагаются в основном в его внешней части и захватывают как биогенную взвесь (фитопланктон и органо-минеральные флоккулы), так и терригенную (минеральную) составляющую. При этом они отфильтровывают частицы (в том числе бактерий) разной величины вплоть до минимальных (с размерами менее 1 мкм).
По имеющимся данным мощность этого биофильтра огромна: весь объем воды из эстуариев крупнейших рек Сибири профильтровывается планктонными рачками за 1–1.5 суток, а весь объем вод Мирового океана – за полгода (Богоров, 1969; Виноградов и др., 1994). В этой же зоне маргинального фильтра обитает большое количество бентосных фильтраторов, также вносящих существенный вклад в процесс потребления вещества, приносимого в море реками. Колония мидий, располагающихся на дне площадью 1 м2, за сутки отфильтровывает взвесь из 100–1000 т морской воды (Кудинова–Пастернак, 1951 и др.).
В общей сложности в эстуариях суммарно за счет физико-химических процессов и активности организмов удаляется около 90% приносимых реками взвешенных веществ и 30% растворенной органики (Лисицин, 2001).
В Белое море впадает множество рек, в том числе и достаточно крупных. В связи с этим роль маргинальных фильтров в эстуариях рек* Кандалакшского, Двинского, Онежского и Мезенского заливов нельзя не учитывать, рассматривая баланс органических веществ.
Специальные исследования показали (Лисицин и др., 2003; Немировская, 2004), что закономерности функционирования маргинальных фильтров, установленные первоначально на основании изучения эстуариев крупных сибирских рек (Лисицин, 1994, 2001), применимы и в данном случае, т.е. при анализе процессов, протекающих в Белом море. Подтверждением этому служат материалы (рис. 8), характеризующие осаждение взвеси в эстуариях двух беломорских рек.
На этом основании можно, исходя из указанных выше соотношений, приблизительно рассчитать, сколько ОВ осаждается в эстуариях и какое количество органики преодолевает маргинальные фильтры беломорских рек и поступает в экосистемы открытого моря.
____________________ * В одной из недавних работ А.Н. Пантюлина (Pantyulin, 2001) было предложено считать все Белое море единым эстуарием, состоящим из четырех (включая само море) эстуарных систем разного масштаба. Обильный речной сток, поступающий в акваторию моря через множество эстуариев, действительно оказывает существенное воздействие на гидрологический режим поверхностных вод Белого моря, но практически не сказывается на характеристиках глубинных вод, имеющих баренцевоморский генезис, высокую соленость и крайне низкие температуры. В этой ситуации едва ли правомочно рассматривать все море в целом как единый эстуарий. Сказанное не означает преуменьшения роли беломорских эстуариев, но, отдавая им должное, рассмотрим участие эстуарных экосистем в общем балансе вещества и энергии позже (гл. 7).
а – р. Северная Двина (27 июня 2002 г.); б – р. Онега (28 июня 2002 г.); в – р. Онега (8 сентября 2002 г.).
Реки, как уже отмечалось, несут в основном взвешенные ОВ (Максимова, 1991). Растворенной органикой, содержащейся в речном стоке, можно без существенной ошибки пренебречь. Взяв за основу уровень осаждения органики (90%) в зоне маргинальных фильтров (Лисицин, 1994, 2003), нетрудно подсчитать, что в эстуариях из 5.5 млн. т ОВ, приносимых беломорскими реками за год, осаждается около 5 млн. т или 2.5 млн. т органического углерода. Значительная часть этих веществ утилизируется обитающими здесь планктонными и бентосными организмами. Остальные захораниваются в осадках и, минерализуясь микроорганизмами, служат в дальнейшем источником биогенных элементов. Та часть принесенных реками взвешенных органических веществ (около 0.5 млн. т или 0.25 млн. т углерода), которая преодолела маргинальные фильтры, разносится течениями по всей акватории Белого моря и включается в баланс вещества и энергии уже в собственно морских экосистемах.
Органические вещества разного происхождения и состава играют неодинаковую роль в энергетических процессах, протекающих в море. Продуцируемые обитателями моря более лабильные ОВ (автохтонная органика), наиболее значимы для различных организмов, поскольку они в гораздо большей степени вовлекаются в трофические процессы. Аллохтонная органика, приносимая реками, утилизируется значительно хуже. О степени утилизации ОВ можно судить, в частности, по отношению биохимического
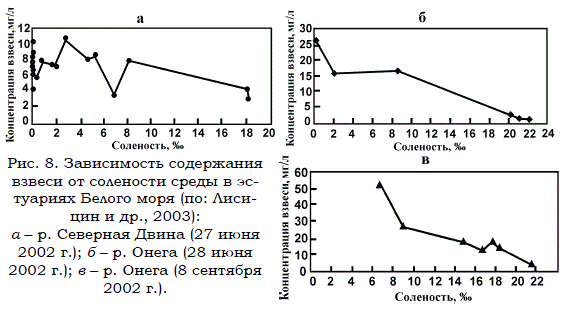
Рис. 8. Зависимость содержания взвеси от солености среды в эстуариях Белого моря (по: Лисицин и др., 2003):
потребления кислорода к перманганатной окисляемости. Для лабильного и хорошо усваемого ОВ фитопланктонного генезиса это отношение близко к единице. Более низкие значения свидетельствуют о большей стойкости и худшей усваимости органики.
По имеющимся данным (Максимова, 1991) в открытых водах Белого моря этот показатель варьирует обычно от 0.5 до 0.75 при средней величине порядка 0.6. В предустьевых пространствах, куда поступает большое количество аллохтонной органики, данное отношение гораздо ниже: около 0.2 (0.15–0.25).
Таким образом, аллохтонная органика, приносимая беломорскими реками, усваивается приблизительно в 3 раза хуже автохтонной. Следовательно, чтобы привести усваимость ОВ различного генезиса к единому знаменателю (усваимости автохтонного ОВ), необходимо общее количество органики речного происхождения (5.5 млн. т) уменьшить в 3 раза, т.е. примерно до 1.8 млн. т (0.9 млн. т органического углерода).
Что касается той части органики (О.5 млн. т), которая, преодолев маргинальные фильтры в эстуариях беломорских рек, проникает в открытое море и разносится течениями по его акватории, то с помощью аналогичной методики можно подсчитать, что она эквивалентна по усваимости примерно 0.2 млн. т автохтонных ОВ.
Таким образом, в эстуариях осаждается и в той или иной степени утилизируется их обитателями или отлагается в грунт количество аллохтонной органики, эквивалентное по усваимости примерно 1.6 млн. т автохтонного ОВ. В пересчете на органический углерод это составляет 0.8 млн. т в год, что эквивалентно 0.8 . 1013 ккал. Та часть взвешенной аллохтонной органики, которая, преодолев маргинальные фильтры эстуариев, проникла в море, содержит около 0.1 млн. т органического углерода, что эквивалентно энергетически 0.1.1013 ккал.
Таким образом, за год в Белое море поступает такое количество аллохтонных органических веществ, которое соответствует по усваимости 1.8 млн. т (0.9 млн. т углерода) автохтонных ОВ и эквивалентно энергетически 0.9.1013 ккал.
Содержание органических веществ, сезонные и пространственные изменения их концентрации и биохимического состава в Белом море более или менее полно начали изучать во второй половине прошлого столетия (Максимова, 1960, 1991; Максимова, Дацко, 1961; Агатова и др., 1994, 2002; Белое море, 1995; Агатова, Торгунова, 1998; Кулаковский, 2000; Agatova et al., 2003).
Общее содержание органических веществ в беломорской воде, согласно имеющимся источникам информации, варьирует на поверхности моря в довольно широких пределах: от 3 до 45 мг/л. Если рассматривать, как концентрация органики в поверхностных слоях воды меняется по сезонам, то оказывается, что ее максимум приходится на июль–август. По мере продвижения на глубину эти сезонные изменения становятся все менее выраженными, а содержание ОВ в воде снижается (Агатова и др., 1994). На глубине 200–300 м содержание органического углерода обычно не превышает 3 мг/л (Максимова, 1960), но в придонном слое воды оно примерно в 1.5 раза выше (4.0–6.3 мг/л), чем в глубинных водах, находящихся над дном в пределах 2–5 м (Агатова и др., 2002).
Самые высокие концентрации органических веществ зарегистрированы вблизи устьев рек, впадающих в Белое море (Максимова, 1991). Прибрежная зона вообще наиболее богата органическими веществами. Одна из самых крупных составляющих баланса органики – органические вещества речного стока и макрофитного происхождения (Максимова, Дацко, 1961; Возжинская, 1986; Невесский и др., 1977; Бек, 1990 а,б).
Среднее содержание органических веществ в воде Белого моря варьирует по данным разных авторов (Максимова, 1960, 1991, 2004; Агатова и др., 1994; Белое море, 1995) от 7 до 19 мг/л. На долю углерода приходится обычно около половины общей концентрации органических веществ (Скопинцев, 1950; Виноградов, 1967). Следовательно, среднее содержание органического углерода во всей толще беломорских вод варьирует от 3.5 до 9.5 мг/л при средней величине около 6.0 мг/л. Это подтверждается и данными, приведенными ниже (табл. 7). Приняв эту величину (6 мг/л) в качестве исходной, можно рассчитать, каково общее содержание органического углерода в воде Белого моря.
Оно составляет приблизительно 3.2.1016 мг, что энергетически эквивалентно 3.2.1014 ккал. При пересчете на столб воды под 1м2 это составит около 3.5.103 ккал.
Результаты этого расчета показывают, насколько велик источник энергии, используемый различными компонентами экосистем Белого моря напрямую или через ряд трансформаций органического вещества.
Основу растворенной органики в водах Белого моря составляют белки, углеводы и липиды (Агатова и др., 1994, 2002). Концентрации аминокислот, регистрируемые в воде Белого моря, примерно в 2 раза превышают аналогичные показатели, обычно приводимые в справочниках при характеристике морской воды (Попов и др., 1979) и специальных работах, посвященных анализу процессов абсорбции аминокислот морскими организмами (Jorgensen, 1979). Данных, позволяющих сравнить Белое и Баренцево моря по содержанию органических веществ, к сожалению, не очень много. Дело в том, что в большинстве случаев исследователи Баренцева моря обращали внимание не на общее содержание органики, а на концентрации отдельных составляющих: липидов, аминокислот, углеводов и др. Тем не менее хотя бы в первом приближении провести такое сравнение возможно.
В августе–сентябре 1976 г. на разрезе от Мурманска к Земле Франца-Иосифа было зарегистрировано содержание растворенного в воде углерода, варьировавшее от 1.0 до 4.8 мг/л, при средней концентрации 2.2 мг/л. Оно уменьшалось с глубиной и по мере продвижения по разрезу с юга на север. Углерод взвешенного вещества в среднем имел концентрацию около 0.08 мг/л (Романкевич и др., 1982). Следовательно, среднее суммарное содержание растворенной и взвешенной органики составляло по углероду около 2.3 мг/л. Если удвоить эту величину, исходя из того, что на долю углерода, как уже отмечалось, приходится обычно около половины массы молекул органических веществ, то получится, что среднее содержание органики в водах исследованной части Баренцева моря составляло приблизительно 4.6 мг/л.
Таблица 7 Содержание (мг/л) органического углерода в воде в разных районах Белого моря (с изменениями по: Аgatova et al., 2003)
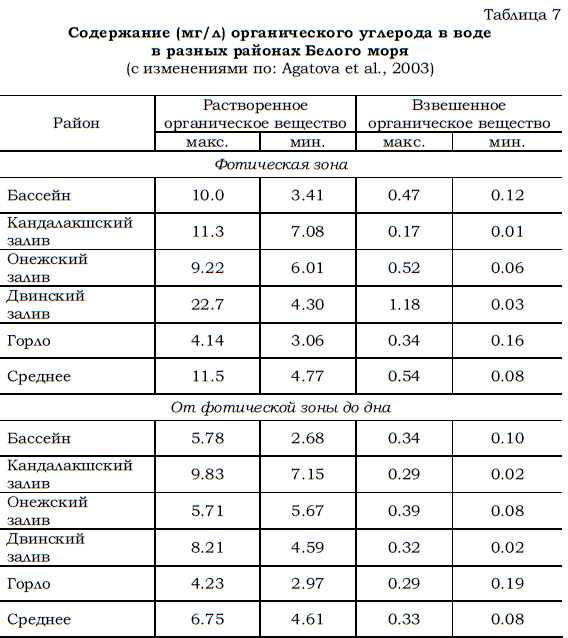
Близкие показатели концентрации органического вещества в воде были получены и в октябре 1998 г. в высокоширотной части Баренцева моря (Агатова и др., 2001). По результатам этих наблюдений установлено, что содержание углерода в растворенных органических веществах варьировало от 1.2 до 5.4 мг/л (табл. 8).
Таблица 8 Содержание органических веществ в водах Баренцева и Белого морей
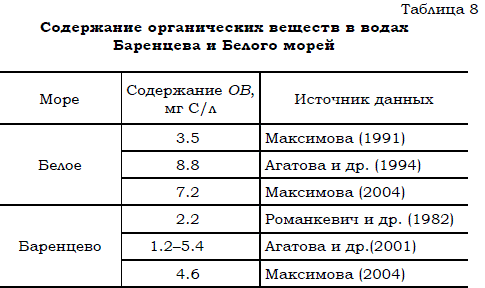
Следовательно, средняя концентрация органики была порядка 6.6 мг/л. При этом, по свидетельству авторов статьи, количество растворенной органики превосходило содержание взвешенной не в 10 раз, как обычно бывает в море (см. выше), а в 30–100 раз.
В связи с этим следует заметить, что соотношение этих форм органических веществ в водах Баренцева моря, рассчитанное по вышеприведенным данным (Романкевич и др., 1982), также было около 30 : 1. Если вспомнить, что растворенные органические вещества имеют в основном автохтонное происхождение, а большая часть взвеси приносится реками, то указанные выше соотношения обеих форм органики становятся легко объяснимыми. В Баренцевом море на долю речного стока приходится лишь очень незначительная часть от общего количества вод, участвующих ежегодно в его водообмене (Максимова, 2004).
Таким образом, сравнение показывает, что средняя концентрация органических веществ в водах Белого моря выше таковой в Баренцевом море.



