ОСНОВНЫЕ ПОНЯТИЯ И МЕТОДЫ ИССЛЕДОВАНИЯ
Биологию принято, в отличие от математики и физики, относить к числу «неточных» наук. В известной степени такая точка зрения имеет основания, хотя и в так называемых «точных» науках положение далеко от идеального. Несмотря на то, что количественные методы анализа приобретают все больший вес, биологами по-прежнему неоднозначно трактуются многие базисные понятия. К ним можно причислить такие, например, как адаптация, популяция, экосистема и многие другие (Мейен, 1990).
В научной литературе слишком часты ситуации, когда одними и теми же терминами обозначаются разные явления, в результате чего исследователи не могут понять друг друга.
В связи с этим, прежде чем начать изложение основного материала, необходимо уточнить применение различных понятий и терминов. При этом автор не стремился к их абсолютизации, хорошо понимая, что такая задача бессмысленна, а попытка ее решения обречена на провал. Мое желание было более скромным, конкретным и вполне осуществимым: придать таким понятиям операционный характер, т.е. указать, как они будут использоваться и каковы границы их применения. При этом будут обсуждаться не все используемые термины, а лишь те из них, трактовка которых неоднозначна. Кроме того, мне представляется излишним приводить ссылки на исследования, в которых те или иные термины использованы ошибочно: достаточно лишь высказать мнение о том, как, на мой взгляд, ими надлежит пользоваться.
Во многих гидробиологических публикациях часто встречаются выражения типа «экосистема моря», «экосистема озера» и т.п., однако такое использование данного термина не соответствует смыслу, который изначально вкладывался в понятие экосистемы. Данный термин был предложен А. Тэнсли (Tensley, 1935) для обозначения системы, состоящей из сообщества организмов (биоценоза) и их биотопа, т.е. комплекса условий обитания и пространственного положения сообщества. Более позднее понятие «биогеоценоз» (Сукачев, 1944), особенно популярное в отечественной геоботанике, не имеет никаких отличий от понятия экосистема, предложенного значительно раньше. По сути дела, это – синонимы. Очевидно также, что в любом водоеме, будь то море, река, озеро или пруд, число биоценозов значительно больше единицы. Соответственно и экосистема не одна; следовательно, и говорить об экосистеме водоема неверно. В соответствии с этим в монографии будут обсуждаться закономерности организации и функционирования отдельных экосистем, а не некой единой экологической системы всего Белого моря.
Биота водоемов, как и биосфера Земли в целом, в организована в виде иерархически соподчиненных систем. Для уровней организации выше экосистемного предложены другие наименования: фации, формации и др., но их обсуждение не входило в нашу задачу. Эти вопросы детально проанализированы в ряде специальных работ (Shelford, 1970; Бабков, Голиков, 1984; Бурковский, 2002, 2006 и др.). На более низких уровнях организации биосистем основное внимание гидробиологов уделяется изучению видов и популяций гидробионтов, и хотя популяция – одна их наиболее исследуемых биологами внутривидовых структур, но по поводу ее определения у специалистов разного научного профиля нет единого мнения. При всем разнообразии трактовок данного понятия наиболее рациональным представляется генетический принцип выделения популяций.
«В генетическом смысле под популяцией понимают сообщество свободно скрещивающихся организмов, с характерным для составляющих ее особей генотипом и фенотипом...» (Лобашев, 1963, с. 425).
Близкое определение приведено в «Большом энциклопедическом словаре. Биология» (2001, с. 499), где написано, что под популяцией принято понимать «...совокупность особей одного вида, обладающих общим генофондом и занимающих определенную территорию».
Аналогично этому в монографии нескольких известных американских биологов (Bush et al., 2001, с. 312) находим следующее высказывание: «Популяция может быть определена как группа организмов одного вида, занимающих в данное время некое единое пространство и имеющих обособленный пул генов» (перевод мой – В.Б.).
Все эти определения едины в том, что популяция представляет собой внутривидовую самовоспроизводящуюся группировку особей, генетически отличающихся от соседних популяций. Строго говоря, до тех пор, пока не доказано, что между рассматриваемыми группами организмов одного вида из тех или иных водоемов (или из их частей) не происходит обмена генами, считать их разными популяциями нельзя. В тех случаях, когда таких специальных исследований не проведено, будет использоваться нейтральный и более адекватный термин поселение (Максимович, 2004).
Вертикальное распределение обитателей моря описывается с помощью ряда специальных терминов, использование которых бывает не всегда однозначным. Значительно различаются, в частности, понятийный аппарат и терминология, используемые гидробиологами, работающими на море или пресных водоемах. В связи с эти кратко охарактеризуем те из них, которые будут использоваться в книге. Зона штормовых заплесков и выбросов получила название супралиторали. Ее нижняя граница соответствует уровню сизигийного прилива.
Ниже следует приливно-отливная зона или литораль, разделяемая на верхний, средний и нижний горизонты. Границы между верхним и средним и между средним и нижним горизонтами литорали определяются соответственно уровнями квадратурных прилива и отлива. Нижняя граница всей литорали и ее нижнего горизонта соответствует уровню сизигийного отлива.
За литоралью следует сублитораль, тянущаяся по вертикали от нуля до глубины 200 м, соответствующей нижней границе континентального шельфа. Ее, в свою очередь, принято делить на несколько зон: адлитораль, циркумлитораль, медиаль, элитораль (Бабков, Голиков, 1984; Луканин и др., 1995; Наумов, 2006 и др.).
Часто особо выделяется фитальная зона, т.е. та часть дна и толщи воды, куда проникает свет и где распространены водные растения. В Белом море нижняя граница фитали соответствует 20 м, хотя в разных частях моря она может смещаться на большие или меньшие глубины в соответствии с прозрачностью воды. Это не означает, однако, что глубже 20 м в толще воды Белого моря нет планктонных микроводорослей. В гораздо меньшем количестве, но они встречаются на этих глубинах (Ильяш и др., 2003), хотя там и нет света, а вот макрофиты, действительно, не проникают глубже 22 м (Денисов, Денисова, 1979).
Зона, простирающаяся по вертикали от 200 м (условная граница континентального шельфа) до 343 м (максимальная глубина Белого моря), получила название псевдобатиали. Беломорская псевдобатиаль представляет собой депрессию дна, не соединяющуюся с батиалью Мирового океана. Это ее отличие от обычной океанической батиали и послужило основанием для специального названия, предложенного А.П. Андрияшевым (1974).
Организмы, населяющие гидросферу Земли, обитают в толще воды, на поверхностной пленке, на грунте и других местах. Обитатели каждого из этих биотопов характеризуются рядом специфических адаптаций. Комплексы организмов, адаптированных к жизни в пелагиали, представлены планктоном и нектоном, на твердых субстратах – бентосом и перифитоном, а на пленке поверхностного натяжения – нейстоном (Зернов, 1934; Константинов, 1986).
Под планктоном понимается «...совокупность совместно обитающих организмов, населяющих водную толщу и недостаточно подвижных, чтобы противостоять перемещению водных масс» (Федоров, 1987; цит. по: Федоров, 2004, с. 347).
Нектонные организмы, также населяющие пелагиаль, отличаются от планктонных тем, что способны гораздо активнее плавать и преодолевать течения. Это не означает, однако, что планктонные организмы не двигаются, а только парят в толще воды. Наоборот, многие из них (например, при вертикальных миграциях) преодолевают большие расстояния: сотни и даже тысячи метров. Величина этих перемещений становится особенно впечатляющей, если вспомнить, что многие планктонные организмы имеют небольшие размеры, обычно не превышающие несколько миллиметров.
В составе планктона имеются организмы разного размера – от микроскопических бактерий, водорослей, простейших и личинок многих животных до некоторых сцифоидных медуз, щупальца которых могут достигать десятков метров в длину. В связи с этим выделяется нано-, пико-, микро-, мезои макропланктон, хотя, насколько мне известно, четких размерных границ между этими группами планктонных организмов не проведено.
Кроме размеров, планктонные организмы принято разделять также по их принадлежности к бактериям, растениям или животным, выделяя соответственно бактерио-, фитои зоопланктон.
Все организмы, обитающие на твердых или мягких грунтах, внутри осадков, на их поверхности или временно связанные с осадками, получили название бентосных. К их числу принадлежат и те, что используют в качестве субстрата как различные искусственные поверхности (сваи, буи, якорные цепи, корпуса судов и т. п.), так и части тела других гидробионтов (например, раковины моллюсков). Их часто выводят за пределы бентоса, выделяя самостоятельную группу организмов-обрастателей. Обсуждение целесообразности такого выделения, а также вопроса о том, относить ли их к бентосным организмам или нет, не входило в задачу автора и увело бы далеко в сторону от рассматриваемых проблем.
В состав бентоса входят бактерии, грибы, растения и животные всех типов. Их размеры различаются на несколько порядков. Для практических целей принято разделять бентос по размерам (Mare, 1942), хотя эта классификация условна, а выделяемые группировки не имеют биологического смысла (Парсонс и др., 1982).
К макробентосу обычно причисляют животных (макрозообентос) и растений (макрофитобентос), размеры которых таковы, что при промывке проб они остаются на сите с ячеей 1 мм2.
Более мелких обитателей дна (от 100 до 1000 мкм) относят к мейобентосу (мезобентосу – Численко, 1961), состоящему из эумейобентоса (фораминифер и мелких многоклеточных организмов – Nematoda, Turbellaria, Harpacticoida, Ostracoda и др.) и псевдомейобентоса (ранних стадий развития макрофауны). Микробентос (1–100 мкм) представлен одноклеточными водорослями (микрофитобентос), протистами (микрозообентос) и микробами (бактериобентос).
В количественных гидробиологических исследованиях принято характеризовать состояние биоты водоема такими показателями обилия, как биомасса и плотность.
Биомасса представляет собой отношение массы организмов к площади дна (если мы имеем дело с бентосными животными и растениями) или к объему воды (если оценивается обилие организмов, обитающих в водной толще). Общая масса организмов того или иного вида на всей акватории или ее части составляет запас.
Количество организмов (численность) всей акватории или ее части, отнесенное к площади дна (бентос) или к объему воды (планктон), именуется плотностью поселения. Этот биологический термин не следует отождествлять с одноименной физической величиной (плотностью).
Под биологической продуктивностью Мирового океана, отдельных морей и других водоемов, а также отдельных экосистем принято понимать способность обитателей того или иного водоема или его части продуцировать органическое вещество (Богоров, 1974). Различия уровня продуктивности оцениваются по величине продукции. Таким образом, продуктивность – способность биоты водоема (или его части) производить некое количество органического вещества, а продукция – количественный показатель продуктивности, характеризующий данное свойство.
Современное понимание термина продукция восходит к известному определению А. Тинемана (Thinemann, 1931): «Продукция видовой популяции за определенное время рассматривается как сумма приростов всех особей, как бывших в наличии к началу изучаемого периода времени, так и вновь отрожденных, включая в величину продукции не только прирост особей, оставшихся к концу периода, но и прирост, который в силу потребления, отмирания и других причин, не входит в конечную биомассу популяции» (Винберг, 1968, с.15).
В этом определении, как подчеркивается далее (с. 16), «...прирост понимается прежде всего как увеличение количества живого или органического вещества данного вида и заключенной в нем энергии (Броцкая, Зенкевич, 1936; Ивлев, 1945 и др.)».
Если исследование показателей обилия (биомассы или плотности поселения/популяции) не предполагает никаких динамических характеристик, то в отличие от них термин продукция, как следует из приведенного определения и общепринятой точки зрения (Одум, 1986; Алимов, 1987, 1989, 1990 и др.), всегда подразумевает время, за которое произошло это накопление. Иными словами, исследуя продукцию, определяют прирост биомассы в единицу времени, т.е. скорость приращения биомассы. При этом исследователи, как правило, интересуются не приростом биомассы отдельного организма, а ее изменениями в поселениях, популяциях или сообществах в целом. Следовательно, изучение продукционных характеристик переводит интересы исследователей на более высокий уровень и позволяет выяснить взаимоотношения отдельных элементов водных экосистем и обосновать рациональное использование биологических ресурсов водоема. Очевидно, что базисным элементом такого подхода является анализ процессов продуцирования и потребления органического вещества на разных уровнях организации биологических систем.
Все организмы существуют за счет поступления энергии извне. Основной источник – солнечная энергия, используемая высшими растениями, водорослями, цианобактериями и пурпурными бактериями для создания органического вещества и построения своего тела путем фотосинтеза. Хемосинтезирующие микрорганизмы (серобактерии, железобактерии, нитрифицирующие бактерии и др.) используют для этих целей энергию химических связей различных неорганических и органических соединений. Свет им для этого не нужен. Фотои хемосинтетики, относимые
к автотрофным организмам, используют для биосинтеза биогенные элементы (углерод, азот, фосфор, кремний и др.), получаемые из воды и (или) грунта.
Продукция автотрофов получила название первичной, а сами организмы относятся к продуцентам. Строго говоря, к продуцентам надо относить не только авто-, но и гетеротрофов, которые также продуцируют вещество и энергию, но делают это не путем фотоили хемосинтеза, а потребляя в пищу другие организмы и (или) растворенную органику. Так или иначе, но на практике название продуценты закрепилось лишь за первичными продуцентами, т.е. за фотои хемосинтетиками.
Они в свою очередь служат источником энергии для гетеротрофных организмов (животных), не способных непосредственно утилизировать солнечную энергию и относимых к числу потребителей (консументов), разделяемых на консументов первого, второго и более высоких порядков. Их продукция именуется вторичной. Передача энергии происходит по пищевым (трофическим) цепям путем поглощения одних организмов другими. Каждая ступень синтеза и утилизации органического вещества называется трофическим уровнем.
Организмов, имеющих единственный источник питания (монофагов), практически не существует. Большинство из них обладает более или менее широким спектром пищи. Вследствие этого они соединены в водоеме не линейными, а разветвленными пищевыми связями, образующими трофические сети.
Роль и положение микроорганизмов (различные бактерии и грибы) в цепи утилизации и передачи энергии и вещества в водоеме невозможно оценить однозначно. Обычно их относят к числу редуцентов, т.е. таких организмов, которые обеспечивают деструкцию взвешенных и растворенных органических веществ, утилизируют отмершие организмы (детрит) и продукты жизнедеятельности живых гидробионтов (различные выделения, фекалии и псевдофекалии). Однако, помимо этого, микроорганизмы служат непосредственно источником пищи для микрозоопланктона. В виде агрегатов они утилизируются также многими другими (более крупными) планктонными и бентосными сестонофагами – организмами, питающимися сестоном.
Сестоном (от греческого сестос – просеянный) принято называть мелкие планктонные организмы и взвешенные в воде неорганические и органические частицы, в том числе и детрит, т.е. все то, что улавливается мелкоячеистой планктонной сетью («Большой энциклопедический словарь. Биология», 2001, с. 571). Кроме того, необходимо отметить, что отмирающие организмы потребляются детритофагами не в чистом виде, а вместе с перерабатывающими их микроорганизмами. Это обстоятельство характеризует детрит даже по определению. В только что упоминавшемся энциклопедическом словаре на с. 175 написано, что «детрит (от латинского детритус – истертый) – мелкие органические частицы (остатки растений, животных, грибов вместе с содержащимися в них бактериями), осевшие на дно водоема или взвешенные в толще воды».
Что касается детрита, то его однозначного определения до сих пор не существует. Не вдаваясь в обсуждение этого вопроса, будем в дальнейшем вслед за А.П. Остапеней (1979, с. 258) под термином детрит понимать «единый комплекс, состоящий из частиц мертвого органического вещества на разных стадиях трансформации и ассоциированных с ним микроорганизмов... Как правило, количество детрита определяется по разнице между общим содержанием взвешенного вещества (сестона) и биомассой живых организмов».
Коэффициент полезного действия продукционных процессов относительно невысокий. На разных уровнях их эффективность может отличаться, но в среднем принято считать, что при переходе с одного трофического уровня на другой около 10% идет на продуцирование органического вещества, а остальные 90% энергии рассеиваются. Показатели обилия и величина продукции при переходе с одного трофического уровня на другой снижаются приблизительно на порядок (Еlton, 1947; Раймонт, 1983 и др.).
Для расчетов продукции тех или иных элементов экосистем используются различные методические приемы, описание которых в этой книге представляется излишним, поскольку они содержатся в ряде хорошо известных публикаций (Винберг, 1960, 1968, 1983; Алимов, 1987, 1989, 1990). Автор в большинстве случаев при расчете продукции тех или иных организмов Белого моря использовал показа
тели скорости оборота органического вещества, представляющие собой отношение продукции (Р) к биомассе (В). Это соотношение вошло в отечественную научную литературу под названием P/B-коэффициента, предложенного Л.А. Зенкевичем (1931). Определенные для организмов, принадлежащих к различным систематическим и (или) экологическим группам, P/B-коэффициенты дают возможность рассчитать продукцию за некое время (час, сутки, год).
В.Е. Заика (1972, с.9) предложил отказаться от этого названия, заменив его термином удельная продукция, под которой понимается «продукция за единицу времени в расчете на единицу массы».
Несмотря на то, что это предложение было сделано более 30 лет назад, но и по сей день большинство гидробиологов пользуется P/B-коэффициентами, а не величинами удельной продукции. Величины P/B-коэффициентов, использованные в книге, были заимствованы мною из литературных источников, указанных по ходу изложения материала.
Иные способы расчета продукции, использованные автором, будут охарактеризованы в соответствующих частях монографии. Расчет потребления вещества и энергии теми или иными организмами производился, исходя из соответствующих литературных данных о коэффициентах усвояемости пищи и эффективности продукции.
К сожалению, многие элементы продукционных процессов в Белом море трудно оценить из-за нехватки необходимых данных, поэтому предпринятую попытку анализа, о котором речь пойдет ниже, следует рассматривать лишь как весьма и весьма приблизительную. Ее результаты еще долго будут уточняться по мере поступления дополнительных материалов. В связи с этим следует, кстати, отметить, что за 12 лет, прошедших с того момента, как такая попытка была впервые осуществлена сотрудниками Беломорской биологической станции Зоологического института РАН (Бергер и др., 1995а,б), расчеты многих количественных характеристик потоков вещества и энергии в Белом море дали результаты, отличающиеся от первоначальных. В данной книге также имеются отличия даже от результатов соответствующего анализа, приведенных в моих последних публикациях.
Ранее (Бергер и др., 1995а, б) в расчетах продукции беломорских организмов, о которых речь пойдет ниже, были использованы величины калорийности тех или иных животных и водорослей, приведенные в работе Е.А. Яблонской (1971), посвященной изучению обитателей южных морей СССР, в основном Азовского и Каспийского. Очевидно, однако, что они, во-первых, устарели, а во-вторых, не все могут быть напрямую использованы как показатели калорийности организмов, обитающих в северных морях. Их употребление могло служить источником ошибок в расчетах. Чтобы свести к минимуму возможность ошибок, все доступные материалы о калорийности морских водорослей, беспозвоночных животных, рыб и млекопитающих, найденные в литературе (Винберг, 19861; Виноградов, Шушкина, 19872 ; Загубиженко, 19693; Карамушко, Карамушко, 19954 ; Коржев, 20045 ; Махмудов, 19666 ; Наумов, Чекунова, 19807; Николаева, 19808; Солдатова, 19809 ; Яблонская, 197110; Petersen, 198511, Klеiber, 197512), были собраны воедино (табл. 1).
При этом без внимания осталась литература, содержащая весьма многочисленные данные о калорийности таких «далеких» от Белого моря объектов, как, например, обитатели тропических районов Мирового океана или различные пресноводные организмы (Шерстюк, 1971; Алимов, 1989 и др.).
Подбор использованных данных производился таким образом, чтобы они касались, во-первых, гидробионтов одних и тех же или близких видов, что и обитающие в Белом море, и, во-вторых, чтобы они относились к водоемам, близким к рассматриваемому морю по биогеографической и гидрологической характеристикам.
Поскольку в ряде случаев авторы работ, сведения из которых использованы в табл. 1, измеряли полученные данные в джоулях, то пришлось перевести их в калории, исходя из соотношения: 1 кал. = 4.2 Дж.
В дальнейших расчетах будут использованы следующие усредненные показатели калорийности (ккал /г влажной массы), рассчитанные на основе данных, приведенных в табл. 1:
– фитопланктон – 0.25, – макрофиты – 0.5, – планктонные Copepoda – 0.5, – бентосные ракообразные – 0.9, – зообентос (без ракообразных) – 0.5, – зообентос (среднее) – 0.7, – сельдь – 1.9, – прочие рыбы (без семги, мойвы и сельди) – 1.1, – тюлени – 4.9.
В книге «Методы определения продукции водных животных» (1968), ставшей для гидробиологов одним из классических руководств, справедливо отмечается, что «...громоздкие цифры общей биомассы или продукции во всем водоеме с трудом поддаются осмыслению и оценке и непригодны для сравнительных целей. Поэтому после расчета величин, относящихся ко всему водоему, эти итоговые данные должны быть отнесены к его площади и выражены в килограммах на 1 га, граммах и калориях на 1м2 и т.д., т.е. в форме, допускающей сопоставление с аналогичными материалами по другим водоемам» (Винберг, 1968, с. 11).
Считая это требование абсолютно справедливым, автор поступал следующим образом: получив данные, характеризующие биомассу или продукцию тех или иных элементов экосистем (планктона, бентоса и т.д.) во всем Белом море, относил их затем к площади и (или) объему моря. Для таких пересчетов были использованы следующие морфометрические характеристики исследуемого водоема (см. гл. 2):
– средняя глубина – 60 м, – площадь – 90 тыс. км2, – объем – 5400 км3.
Таблица 1 Калорийность гидробионтов*
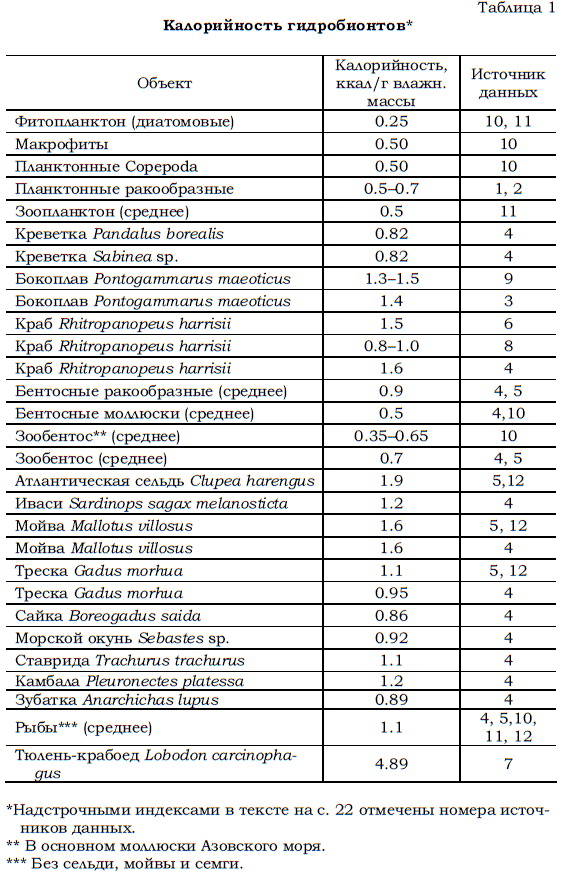
*Надстрочными индексами в тексте на с. 22 отмечены номера источ
ников данных. ** В основном моллюски Азовского моря. *** Без сельди, мойвы и семги.



