СОВРЕМЕННЫЕ ТЕНДЕНЦИИ ИЗМЕНЕНИЙ ПРЕСНОВОДНЫХ ЭКОСИСТЕМ ЕВРО-АРКТИЧЕСКОГО РЕГИОНА
Поверхностные воды занимают важнейшее место среди природных ресурсов Арктики. Озера и реки во многом определяют экономическое и социальное развитие северных регионов, тесным образом связаны с культурным наследием коренных народов, являются составной частью их жизненной среды. Пресные воды служат источниками водоснабжения населения и промышленных объектов, источниками энергии, транспортными артериями многих регионов, объектами коммерческого и спортивного рыболовства, рыбоводства, рекреационными объектами, способствующими развитию туризма.
В то же время водные экосистемы являются наиболее уязвимыми компонентами природной среды Арктики, поскольку интегрируют все изменения окружающей среды, происходящие на территории их водосборов. Вследствие особенностей распространения воздушных масс Северного полушария Земли большая часть загрязняющих веществ, выбрасываемых в атмосферу в индустриально развитых регионах, переносится в Арктику, где они осаждаются и накапливаются в экосистемах. Значительна роль и местных источников загрязнения.
Существенную роль в изменении условий обитания в арктических регионах играют изменения климата, имеющие глобальный характер. И хотя общая направленность таких изменений не однозначна, а в отдельных районах может иметь разнонаправленный характер, не вызывает сомнения разбалансировка климатической системы, снижение стабильности, увеличение
ее, по образному выражению академика Ю.И. Израэля, «нервозности». В последние годы в высоких широтах регистрируются наибольшие уровни проявления признаков глобальных и региональных преобразований климатических процессов, включая аномальные отклонения температуры на фоне тренда на потепление (Polyakov et al., 2002; McBean et al., 2005; ACIA, 2005). Реальное влияние связанных с изменением климата процессов показывает высокую сложность этих взаимодействий, особенно во взаимосвязи с многофакторной антропогенной нагрузкой. Многогранность
возможных реакций на происходящие процессы со стороны экосистем при возможности задержки, их перекрывания, проявления синергетических, аддитивных и антагонистических механизмов делает крайне сложными попытки предсказания возможных последствий.
Проблема оценки и прогноза степени деградации водоемов под воздействием антропогенных загрязнений и глобальных изменений окружающей среды остается по-прежнему актуальной. Не реально осуществлять контроль за всеми абиотическими и биотическими параметрами даже самых простых пресноводных экосистем. Для оценки их состояния используется ограниченное количество доступных показателей, которые можно разбить на две основные группы: определение уровней веществ-загрязнителей в воде и/или в седиментах и использование биомаркеров (включая определение уровне веществ-загрязнителей в биоте). Каждый метод имеет специфические преимущества и недостатки. Биологический эффект того или иного вещества не может быть строго детерминирован лишь его концентрацией в водной среде. Эта зависимость носит вероятностный характер и определение токсического воздействия веществ-загрязнителей на биоту водоема лишь по их количественным показателям в воде или донных отложениях и сравнение их со стандартизированными показателями качества среды (например, ПДК) имеет весьма относительную экологическую ценность.
Биологические показатели состояния экосистем (видовое разнообразие, популяционные и организменные показатели и т.д.) имеют очевидные преимущества, так как являются "интегрирующими" показателями всех изменений за продолжительный период времени и непосредственно отражают ответы биоты на тот или иной вид антропогенного воздействия. Однако выявить и интерпретировать последствия, обусловленные антропогенным фактором, довольно сложно. Показатели сообщества и популяций часто опосредованы процессами, происходящими на более низких трофических уровнях, и протекают на фоне их естественных колебаний. Понятие "нормальные показатели" для биологических систем довольно условно и непостоянно. Оно зависит от региональных, сезонных, меж- и внутривидовых, половых различий, физиологического состояния организма и колебаний нормальных факторов внешней среды и способности организмов к ним приспосабливаться. Кроме того, трудности в интерпретации наблюдаемых эффектов обусловлены неспецифичностью ответов биологических систем высокого уровня. Поэтому проведение лишь биологических исследований, без комплексного анализа качества окружающей среды не может дать ответы на все вопросы.
Необходимо также добавить ряд специфических региональных особенностей, осложняющих интерпретацию получаемых результатов исследований водоемов Евро-Арктического региона. Как правило, загрязнение сточными водами носит относительно локальный характер, в то время как
остальные территории подвергаются аэротехногенному типу загрязнения, для которого свойственна следующая специфика:
• нагрузка носит специфический характер, обусловленный долговременной и сезонной динамикой выпадения загрязнителей из атмосферы: наблюдается пиковое увеличение нагрузки токсикантов в короткие периоды снеготаяния или обильных дождей на фоне общего медленного возрастания их концентрации за относительно продолжительный период;
• постепенное накопление в наземных и пресноводных экосистемах, возрастание общего количества долгоживущих загрязняющих веществ, циркулирующих в этих экосистемах;
• скорость миграции загрязняющих веществ в наземных и водных экосистемах обусловлена геологией водосборов, ландшафтным характеристиками, морфологией водоемов и другими факторами;
• большая часть загрязняющих веществ, выпадающих на территорию водосбора, в конечном итоге оказывается в водоеме, и их состояние может быть показателем нагрузки на весь этот район.
В целом можно выделить следующие региональные особенности, которые необходимо учитывать при изучении водоемов Севера.
• Природная вариабельность параметров природных сред и биологических систем. Для региона характерно чрезвычайно высокое разнообразие типов ландшафтов, что затрудняет проведение сравнительных исследований, так как даже на небольшом участке могут оказаться водоемы с радикально отличающимися природными условиями. Отсутствие четких представлений о внутриводоемных процессах, включая определение токсичности и биодоступности загрязняющих веществ для гидробионтов. Особенности арктических пресноводных экосистем (короткий вегетационный период, короткие пищевые цепи, высокая эффективность утилизации доступной энергии, уязвимость структуры сообществ и зависимость ее от отдельных видов).
• Особенности распространения, выпадения, миграций, трансформации загрязняющих веществ. Специфика процессов глобального и локального загрязнения окружающей среды, на фоне низкой минерализации и буферной емкости поверхностных вод. Отсутствие четких границ начала регионально-фоновой зоны, что нередко приводит к методической ошибке выбора контрольных объектов, связанных с оценкой интенсивности и степени воздействия на объект, расположенный в локальной зоне источника загрязнения. Отсутствие объективных показателей нагрузки на водоем за продолжительный период времени: до сих пор не создано надежных методов количественной оценки поступления загрязняющих веществ в различные компоненты природной среды за долговременную ретроспективу. Причем определение дозы важно не только для зоны локального загрязнения, но также и регионально-фоновой зоны.
• Аккумуляция долгоживущих загрязняющих веществ в экосистемах, постоянное увеличение их общего количества, циркулирующего в экосистеме. Существуют трудности аналитического определения долгосрочных изменений их низких уровней в природных средах. Отсутствие достоверной информации о состоянии экосистем в «доиндустриальный период». Отсутствие достоверных долговременных рядов наблюдений. Неопределенность и неспецифичность ответных реакций биологических систем на продолжительное воздействие малых сублетальных доз.
Трудности определения биологической опасности того или иного вида загрязнения можно наглядно продемонстрировать на примере загрязнения окружающей среды тяжелыми металлами, приоритетными поллютантами региона. Хорошо известно, что тяжелые металлы являются наиболее опасными загрязняющими веществами, поступающими в окружающую среду в результате человеческой деятельности из-за их высокой токсичности и потенциальной биоаккумуляции в живых организмах, в том числе и гидробионтах. В то же время при оценке опасности загрязнения нельзя руководствоваться только количественными показателями содержания того или иного вещества. Его токсичность и биодоступность в природном водоеме будет определяться множеством абиотических и биотических факторов. Экотоксикологический эффект действия тяжелых металлов зависит от биологической особенности объекта, качества среды, типов соединений, в которых находится металл, продолжительности воздействия, путей поступления металла (через воду, донные осадки, пищу), физиологического состояния рыб, эффектов взаимодействия с другими стрессовыми факторами, в том числе и с другими металлами, способности рыб перемещаться в незагрязненные участки и др. Все эти факторы могут напрямую или косвенно зависеть от рН, температуры и других физических и химических факторов.
Кислотность среды является основным фактором, контролирующим и регулирующим вариабельность во многих химических, биологических и геологических процессах. Понижение рН является причиной многих дополнительных изменений качества воды, потенциально воздействующих на водные организмы. Важнейший эффект рН - влияние на растворимость и формы нахождения токсичных металлов. Концентрация ионов металла, которые являются более биодоступными формами элементов, как правило, увеличивается со снижением рН.
Хотя выбросы кислотообразующих соединений предприятиями цветной металлургии региона в последние годы сократились, они по-прежнему остаются значительными, обусловливая процессы закисления поверхностных вод в кислоточувствительных районах и вызывая весенние рН-шоки практически по всей территории региона. При этом в последние десятилетия выбросы в атмосферу тяжелых металлов практически не сокращаются (рис.1).
Помимо кислотности на биодоступность металлов оказывают влияние и другие факторы среды. Так, в противоположность низким рН, ионы Са2 и Mg+2 могут проявлять защитное действие от тяжелых металлов, которое заключается в их успешной конкуренции с последними на поверхности жабр рыб. Многие природные органические соединения образуют с тяжелыми металлами малорастворимые комплексы. В естественных водоемах вещество-загрязнитель или какая-то его часть может находиться в связанных формах (комплексах или ассоциациях), что делает его полностью или частично недоступным для биоты. При этом вещество-загрязнитель не оказывает прямого токсического воздействия на водные экосистемы, но может рассматриваться как потенциальная угроза окружающей среде, если возможно изменение его биодоступности под воздействием каких-либо дополнительных факторов (например, изменение рН). Необходимо также учитывать «постоянство» токсиканта (т.е. его время присутствия в водоеме) и риск бионакопления в пищевых цепях, что может вызывать увеличение концентрации токсиканта в организмах высших трофических уровней. Связанные элементы могут быть абсорбированы микроорганизмами и включены в пищевые цепи или непосредственно попадают в организмы животных более высоких трофических уровней с пищей (фильтраторы, бентофаги и т.д.).
Рис.1. Динамика выбросов в атмосферу загрязняющих веществ ОАО «Кольская горно-металлургическая компания» (http://www.kolagmk.ru/ecology/monitoring)
Таким образом, токсичность и биодоступность тяжелых металлов в природных водоемах зависит от очень большого числа факторов. При оценке антропогенных изменений состояния экосистемы перед исследователями всегда стоит проблема выбора ее характеристик, адекватно отражающих протекающие процессы.
Многолетние комплексные исследования водоемов Севера Европы, выполненные Лабораторией водных экосистем ИППЭС КНЦ РАН, позволяют сделать вывод, что показатели состояния сообществ гидробионтов объективно отражают состояние окружающей среды и могут быть использованы для ее оценки. Сравнительное исследование биологических систем различного уровня организации в условиях, как естественных, так и в антропогенно-измененных, позволяет непосредственно выявить эффекты, обусловленные воздействием загрязняющих веществ за продолжительный период времени, определить пороговые уровни нагрузок и дать наиболее реалистичный прогноз развития. Результаты исследований позволяют выявить ряд закономерностей в изменениях структурно-функциональной организации популяций гидробионтов, а также специфические изменения в их организмах в условиях загрязнения водоема тяжелыми металлами. Установлены определенные закономерности и зависимости наблюдаемых эффектов от величины нагрузки. Качественные и количественные показатели могут быть использованы для биоиндикации данного вида загрязнений и определения критических нагрузок.
Тенденции изменений водорослевых сообществ
Важнейшим элементом водных экосистем являются первичные продуценты, основными из которых в северных областях являются водоросли. Различные показатели состояния сообществ водорослей являются признанной обязательной составляющей системы экологического мониторинга (Баринова, Медведева, 1996; Комулайнен, 2003, 2007; Баринова и др., 2006; Комулайнен и др., 2006;). Вместе с тем, изученность пресноводных водорослевых сообществ региона, особенно в «доиндустриальный» период и период наибольшей индустриальной активности, остается недостаточной. Наиболее ценные сведения о водорослях систематизированы в ряде работ (Комулайнен и др., 2006). Имеются данные первых флористических исследований водорослей на Кольском полуострове (^аЫапЬещ, 1812; Ку1ап<іег, 8ае1ап, 1859; КіШтап, 1890; КіШтап, Раїтеп, 1889). Большой вклад в изучение состава и региональных особенностей альгофлоры, а также последствий антропогенных преобразований экосистем субарктических водоемов внесли сотрудники Института озероведения РАН (Давыдова, 1971; Летанская, 1974) и Карельского научного центра (Комулайнен, 2003; 2007). В последние годы получены данные о видовом составе и структуре водорослевых сообществ Кольского Севера (Каган, Денисов, 2002; Шаров, 2004; Денисов, Кашулин, 2007; Денисов, 2007; 2009; 2010; Кашулин и др., 2008; 2009). Тем не менее, информация о водорослях многочисленных разнотипных малых озер региона практически отсутствует или является отрывочной. Многие результаты получены по данным разового отбора проб, поэтому могут не отражать реальное состояние водоема, особенно это относится к фитопланктону, численность и видовой состав которого способны к резким изменениям в ответ на меняющиеся внешние условия, что особенно ярко проявляется в малых озерах. В этих обстоятельствах велика роль диатомового анализа донных отложений, позволяющего реконструировать динамику интегральных параметров, отражающих состояние и развитие экосистемы в историческом прошлом.
Анализ имеющихся данных позволяет сделать выводы об основных тенденциях изменений, происходящих в сообществах водорослей за последние десятилетия. Происходят радикальные перестройки структурно-функциональных характеристик альгоценозов, изменение направления и скорости сукцессий как результат взаимозависимого действия целого комплекса регулирующих факторов, из которых наиболее значимыми являются антропогенные, в настоящее время усиленные аномальной динамикой региональной климатической системы (рис.2).
Рис.2. Схема трансформации водорослевых сообществ и определяющие этот
процесс факторы
Водорослевые сообщества чутко реагируют на все происходящие изменения: формируются новые адаптационные механизмы, меняются
продукционные характеристики сообществ и экологические предпочтения отдельных видов. Это приобретает определяющее значение для биоиндикации качества среды и задач гидроэкологического мониторинга. При оценке качества вод на основе гидробиологических показателей в настоящее время требуется учитывать синергетику нового комплекса факторов, действие которых неоднозначно и не всегда может быть оценено с помощью существующих общепринятых комплексных показателей. Очевидно, ряд критериев оценки качества среды на основе водорослевых сообществ, требует корректировки и пересмотра для получения объективных результатов биоиндикации состояния водных экосистем.
Изменение степени сапробности. Сапробность представляет собой способность организма выживать в условиях так называемого «органического» загрязнения, под которым понимается поступление элементов минерального питания (фосфаты и нитраты) растительных организмов, продуцентов органического вещества, а также гуминовых веществ и органических соединений различного происхождения. Как правило, загрязнение этого типа сопряжено с развитием процессов эвтрофирования. В рамках реализации мониторинга состояния окружающей среды, проведения инженерно-экологических изысканий, процедуры ОВОС и др. для оценки качества вод по гидробиологическим показателям используются нормативы, закрепленные ГОСТ 17.1.3.07-82. Для определения качества вод и степени их загрязнения рекомендован индекс сапробности (5), выражающий степень загрязнения вод (рис.3) (Сладечек, 1967; Баринова и др., 2006; 1986; 8^есек, 1973).
глобальные антропогенные преобразования окружающей среды • „1
глобальные изменения окружающей среды и климата
природные процессы • Ч.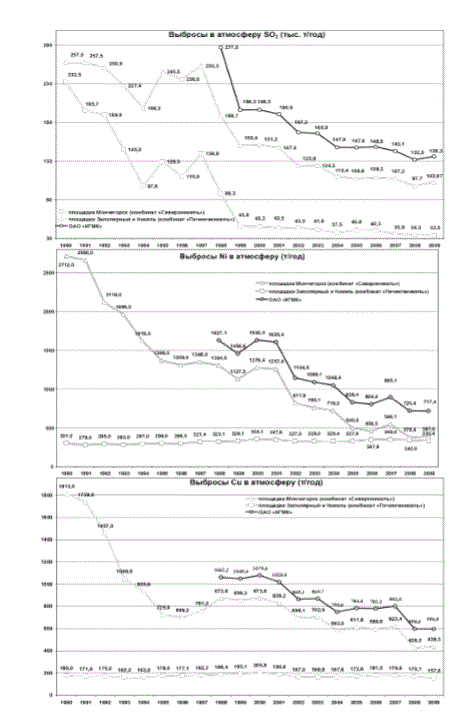
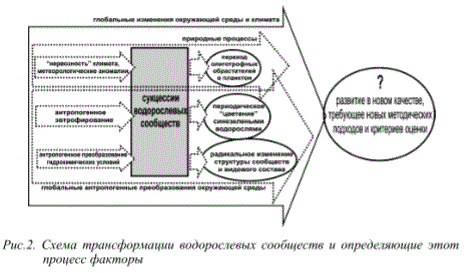

Рис.3. Классификация качества вод и соответствующие степени сапробности, характеризующие процессы эвтрофикации и самоочищения (ГОСТ 17.1.3.07-82)
Реакция водорослей на органическое загрязнение является сравнительно хорошо изученной областью, что позволяет наиболее адекватно оценивать качество вод на основе сапробного индекса, рассчитанного по альгоценозам. В то же время индекс сапробности представляет собой интегральный показатель состояния альгоценозов и отражает не только степень органического загрязнения, но и особенности внутренней организации водорослевых сообществ, межвидового конкурентного взаимодействия, а также косвенно характеризует биотопические характеристики.
Первоначально индекс сапробности был разработан для оценки уровня органического загрязнения и развития процессов эвтрофикации и не учитывал возможности одновременного воздействия целого комплекса различных факторов, в том числе токсического воздействия и новых сезонных вариаций аномалий метеорологических условий.
К настоящему времени обобщены и систематизированы данные о состоянии водорослей планктона и перифитона водоемов различных ландшафтов Кольского полуострова. В не нарушенных антропогенной деятельностью водных объектах структурно-функциональная организация водорослевых сообществ определяется локальной дифференциацией условий в пределах ландшафтного комплекса. Все воды относятся к лимносапробной категории $ от 0 до 2.0) и характеризуются ультраолиготрофным и олиготрофным трофическим статусом по содержанию хлорофилла «а». Фитоперифитон изученных водоемов отличается большим числом видов, численностью и биомассой (по сравнению с фитопланктоном) и зачастую является основным источником фитомассы, образуемой автотрофами в водоемах, что особенно характерно для мелководных тундровых озер (рис. 4).
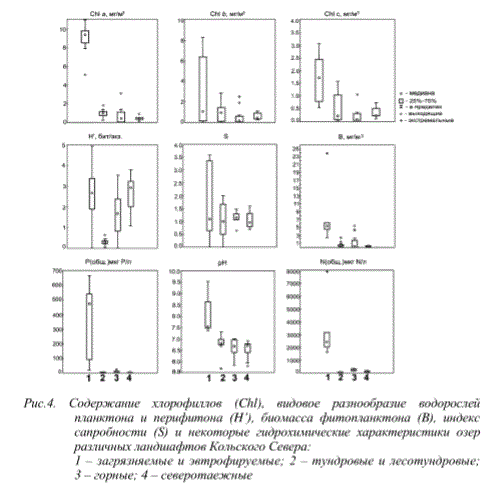
Рис.4. Содержание хлорофиллов (Chl), видовое разнообразие водорослей планктона и перифитона (H’), биомасса фитопланктона (B), индекс сапробности (S) и некоторые гидрохимические характеристики озер различных ландшафтов Кольского Севера:
1 - загрязняемые и эвтрофируемые; 2 - тундровые и лесотундровые; 3 - горные; 4 - северотаежные
Показано, что наибольшими значениями сапробного индекса, отражающего нагрузку биогенными элементами (S), характеризуются водные объекты, испытывающие загрязнение предприятиями по добыче и переработке апатитовых руд (ОАО «Апатит» и Ковдорский ГОК), а также подверженные антропогенному эвтрофированию (стоками очистных сооружений), что обусловлено экстремально высокими для вод Кольского Севера концентрациями N и P. Повышенные значения pH также стимулируют развитие водорослей (рис.4). Водоемы этой группы различаются набором антропогенных факторов и их сочетанием, что отражается на водорослевых сообществах широким диапазоном значений индекса видового разнообразия и сапробности. В то же время водоемы, испытывающие токсическую нагрузку или характеризующиеся высоким содержанием минеральной взвеси, отличаются сравнительно небольшими значениями S, на уровне олигосапробных вод (0.8-1.2) в силу угнетения развития водорослей - сапробионтов. Вероятно, для подобных случаев необходимы корректировка сапробного индекса и его адаптация для оценки качества вод по принятым в РФ нормативам либо ограничения по его применению (ГОСТ 17.1.3.07-82).
Усиление эвтрофикации. Долговременная эвтрофикация вод и интенсификация этих процессов в последние годы проявляется не только в условиях загрязнения стоками предприятий апатитовой промышленности и очистных сооружений городов, но и приобретает глобальный характер и регистрируется в различных районах Кольского полуострова. Степень вклада антропогенных факторов в процессы эвтрофикации вод наглядно демонстрируют величины содержания хлорофилла «а» и максимальной биомассы водорослей планктона. Современные тенденции изменения трофического статуса вод характеризуются значительным усилением скорости этих процессов, а также сильным отклонением ряда показателей от средних значений (рис.4). На рис.5 показана многолетняя динамика значений биомассы фитопланктона для двух участков акватории оз.Имандра и озерно-речной системы Пасвик.
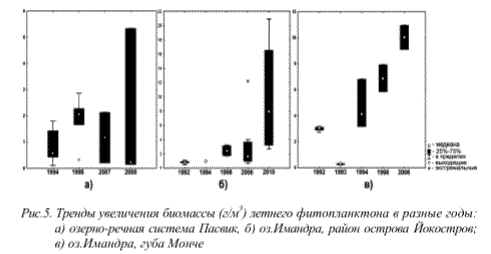
Рис. 5. Тренды увеличения биомассы (г/м3) летнего фитопланктона в разные годы: а) озерно-речная система Пасвик, б) оз.Имандра, район острова Йокостров; в) оз.Имандра, губа Монче
Из этих данных видно, что в последние годы увеличиваются как средние значения биомассы фитопланктона (для оз.Имандра), так и их диапазон - больше экстремальных значений. Это является свидетельством кратковременных периодов массового развития фитопланктона (вплоть до «цветения» воды) в отдельные сезоны. Следует отметить, что уровень биогенной нагрузки антропогенного происхождения в последние годы сохранялся примерно на одном уровне, в то время как значения биомассы (с учетом экстремальных) выросли в несколько раз. Очевидно, процессы эвтрофирования стали усиливаться другими факторами, из которых наиболее мощным представляется глобальное изменение климата в сторону потепления.
В последние годы (2006-2010) в водоемах различного типа Мурманской области явления массового развития водорослей планктона, вплоть до цианопрокариотического цветения воды, наблюдаются регулярно.
Исследования сезонной динамики сообществ фитопланктона, проведенные на базе комплексного мониторингового полигона «Имандра», зарегистрировали периоды массового развития водорослей. Наиболее интенсивное цветение вод наблюдалось в мелководных прогреваемых заливах в районе острова Большой Йокостров. Основными массовыми видами летнего фитопланктона (июль-август) являются перидинеевые (Ceratium hirundinella (O.F. Mull.) Bergh f. hirundinella; Peridinium goslaviense Wolosz) и синезеленые водоросли (Anabaena lemmermannii P. Richt.; A. flos-aquae Born. et Flah.
f flos-aquae). При этом цианобактерии развиваются эпизодически в условиях штиля в заливах, а перидинеи - в открытых участках акватории. Массовое развитие перидинеевых водорослей в штилевую погоду придает поверхности вод рыжевато-красный оттенок. Во время массового развития синезеленых водорослей биомасса может достигать экстремально высоких значений - свыше 21 г/м3. Развитие цианей приводит к изменению гидрохимических характеристик, оптических и органолептических свойств воды. К этим же периодам приурочена гибель молоди сиговых рыб. Хорошо известно, что выявленные в этот период в озере массовые виды Cyanoprokaryota способны выделять в воду токсичные для гидробионтов вещества (цианотоксины), что и могло послужить причиной гибели молоди рыб.
Изменение продукционньх процессов. Являясь фотоавтотрофами, водоросли формируют первичную биомассу, служащую основой для функционирования последующих трофических уровней экосистемы. Скорость образования первичной продукции и уровень образуемой биомассы представляет собой фундаментальную характеристику состояния экосистемы в целом. Изменчивость процесса продуцирования экосистем в сезонном или иных аспектах, а также под влиянием всей антропогенной деятельности позволяет сделать вывод о том, что процесс продуцирования является основным источником эволюции экологических процессов. Важность такого подхода к продуцированию очевидна, поскольку изменчивость экосистем и слагающих их объектов с присущими им функциональными особенностями приводит к возникновению нового уровня адаптации и является регулятором устойчивости экосистемы. В условиях Субарктики скорости процесса первичной продукции и уровень образуемой биомассы были невелики (Летанская, 1974). С появлением новых антропогенных факторов и развитием процессов эвтрофикации вод величины первичной продукции многократно возросли. Максимальные значения биомассы фитопланктона в эвтрофируемых водах могут на несколько порядков превышать «фоновый» уровень (рис.4).
До недавнего времени считалось, что скорость продукционных процессов в олиготрофных водоемах, не испытывающих непосредственного загрязнения элементами питания растительных организмов, остается в пределах условно-«фоновых» значений. Особенно это справедливо для малых горных холодноводных водоемов. Тем не менее, в последние годы были зарегистрированы не типичные для региона явления массового развития водорослей для водоемов, развивающихся в условиях отсутствия прямого антропогенного загрязнения. В малых олиготрофных горно-тундровых озерах Хибинского горного массива в 2010 г. был отмечен период массового развития нитчатых зеленых водорослей (Zygnema sp., Spirogyra sp., Draparnaldia plumosa (Vauch.) Ag.) в толще воды. Данное явление отмечалось даже в маленьком (<0.1 км2) оз.Каровое, расположено на высоте около 410 м над уровнем моря. В оз.Малый Вудъявр массовое развитие нитчатых зеленых водорослей приводило к засорению рыболовных сетей при ихтиологических работах.
Необходимо отметить, что переход представителей перифитона в планктон традиционно считается признаком эвтрофикации водоема. В данном случае речь идет о типичных представителях холодноводного перифитона горных рек, развивающихся в условиях низкого содержания биогенных элементов и высокого содержания кислорода. Эти же водоросли населяют питающие озеро реки. Причины перехода в планктон требуют дополнительных исследований. Вероятно, осеннее перемешивание воды и сочетание метеорологических условий сделало возможным массовое развитие именно этих, не требовательным к биогенным
элементам водорослей. Очевидно, сам этот факт следует рассматривать как индикатор глобальных изменений в экосистемах региона, проявляющихся в том числе и в «нервозности» метеорологических факторов в течение сезона.
Потенциальную возможность такого массового развития водорослей в горных олиготрофных водоемах в конце периода вегетации следует учитывать при проведении сезонных исследований. Очевидно, необходимо поставить вопрос о продукционных возможностях олиготрофных водоемов в отдельные периоды, в качестве основы для последующих прогнозов трансформаций субарктических водоемов под влиянием глобальных изменений окружающей природной среды.
Изменение таксономической структуры сообществ. О долговременных изменениях таксономического состава и соотношения видов в сообществах водорослей в региональном масштабе можно судить на основе информации, полученной в различные периоды исследований, систематизированных в работах авторов (Яковлев, 1991; Комулайнен и др., 2006), а также по результатам диатомового анализа донных отложений (Порецкий и др., 1934). Некоторые сведения из ранних исследований водоемов флористического направления содержат ценную информацию о видовом составе водорослевых сообществ из тех районов Кольского Севера, которые и по сей день являются слабо изученными в альгологическом отношении (Зинова, Нагель, 1935; Жадин, Герд, 1961; Нилова, 1966). Достаточно полной и подробной информацией в настоящее время обладают издания, посвященные крупным озерам (Озера различных ландшафтов ..., 1974; Большие озера ..., 1976; Шаров, 2004). На основе этих данных, а также базируясь на результатах последних исследований, можно заключить следующее. Характер устойчивых тенденций в долговременных изменениях водорослевых сообществ определяется локальным действием группы наиболее мощных регулирующих факторов для конкретного водного объекта, например антропогенных. Так, для различных участков акватории оз.Имандра наблюдаются различные изменения в видовой структуре сообществ водорослей в зависимости от вида и интенсивности антропогенного воздействия. В удаленных от промышленных предприятий водоемах регистрируются изменения, связанные с глобальным загрязнением атмосферы. Последствия такого воздействия для водной экосистемы определяются гидролого-геохимическими условиями формирования вод на водосборе, а потому могут привести к различным сценариям изменения таксономической структуры водорослей для каждого озера или водотока. При этом экстраполяция выявленных закономерностей на весь регион представляется весьма затруднительной, в первую очередь, в силу следующих причин:
1) высокое многообразие разнотипных водных объектов на территории Кольского полуострова;
2) огромное количество регулирующих таксономическую структуру сообществ факторов, как природных, так и связанных с антропогенной деятельностью;
3) недостаточная изученность причинно-следственных связей в системе процессов, определяющих направление сукцессий;
4) недостаток и (или) отсутствие информации об исторической динамике видового состава в прошлом.
Тем не менее, несмотря на всю сложность, многообразие и разнонаправленность хода сукцессионных процессов пресноводных альгоценозов на Кольском Севере, можно обозначить некоторые общие закономерности в изменениях таксономической структуры сообществ. Во-первых, наиболее выраженные
изменения являются прямым следствием долговременной антропогенной нагрузки, связанной как с локальными источниками, так и с глобальным аэротехногенным загрязнением. Во-вторых, таксономический состав чувствителен к изменениям окружающей природной среды и климата, действие которых, в свою очередь, накладывается на комплекс антропогенных факторов.
Облик альгофлоры водных объектов до начала интенсивного промышленного загрязнения характеризовался доминированием в планктоне и перифитоне диатомовых, зеленых (из которых массовыми являлись десмидиевые) и золотистых водорослей. Это были типично олиготрофные и ультраолиготрофные сообщества, в которых абсолютное большинство видов развивается в условиях низкой минерализации. Для водоемов северотаежных природно-ландшафтных комплексов в составе альгоценозов присутствовал ацидофильный комплекс видов (предпочитающий pH <7.0), как результат влияния болотного подкисления и присутствия гуминовых кислот в воде. В перифитоне малых рек доминирующими по численности и биомассе группами были виды рода Zygnema, Oedogonium, в зоне северной тайги - красные водоросли рода Batrachospermum.
Добыча и переработка минерального сырья и строительство городов повлекли за собой возникновение мощного антропогенного пресса на целый ряд водных объектов. Наиболее значимым фактором для водорослей явились процессы эвтрофикации. В альгоценозах это проявилось снижением доли золотистых, для зеленых отмечено сокращение доли десмидиевых водорослей, с одновременным увеличением вольвоксовых, а также усилением роли семейства фрагилариевых в сообществах диатомовых водорослей. При этом в водоемах, подверженных непосредственному загрязнению и эвтрофикации, снижается группа ацидофилов, а в некоторых удаленных от промышленных территорий водных объектах, напротив, доля ацидофилов увеличивается, как результат выпадения кислотообразующих соединений. Изменение гидрохимических условий и рост общей минерализации вод привели к сокращению численности галофобов и увеличению доли галофилов, а в некоторых водоемах - мезогалобов. В перифитоне рек, подверженных непосредственному антропогенному загрязнению, изменение таксономического состава проявилось в увеличении доли улотриксовых, а также диатомовых сем. Fragilariaceae.
В последние годы действие антропогенных факторов, очевидно, было усилено за счет естественной динамики регионального и глобального климата. Это проявилось в первую очередь в резких и кратковременных сезонных изменениях таксономической структуры, связанных с массовым развитием отдельных видов. В составе альгоценозов увеличилась доля синезеленых водорослей, включая и токсичные формы. Периодически наблюдаются этапы массового развития динофитовых водорослей, причем не исключено, что в водах с высоким трофическим статусом они частично переходят на гетеротрофное питание. В водоемах «фоновых» районов Кольского Севера в настоящее время сохраняется типично «субарктический» облик альгофлоры.
Таким образом, мощное антропогенное загрязнение и эвтрофирование вод на фоне глобальных климатических изменений привело к изменению структуры и количественных показателей водорослевых сообществ субарктических водоемов различных ландшафтов. Установлены тенденции резкого увеличения скорости процессов эвтрофикации вод в последние годы, что проявляется в различных районах Кольского полуострова. Значительное ускорение темпов преобразования структурно-функциональных показателей водорослевых сообществ подтверждается палеоэкологическим исследованиями диатомовых комплексов современных донных отложений озер. Так, для малых
горных водоемов Хибинского горного массива, развивающихся в условиях «фона», видовой состав и структура диатомовых комплексов верхних слоев отложений (что отражает современное состояние водоема) имеет существенные отличия от более глубоких слоев. Следовательно, существенные преобразования происходят именно на современном этапе и затрагивают последние десятилетия. Сочетание нагрузки биогенными элементами с токсическим загрязнением для ряда водоемов приводит к искажению таких показателей, как индекс сапробности (S), что заставляет поставить вопрос о разработке корректировок расчетов сапробного индекса для подобных случаев. Наблюдаются периодические вспышки численности токсичных синезеленых водорослей, сопровождающиеся гибелью молоди сиговых рыб. Впервые для Кольского полуострова были зарегистрированы явления перехода в планктонное состояние представителей олиготрофного перифитона в малых горно-тундровых водоемах Хибинского горного массива, что требует переоценки продукционных характеристик озер этого типа и пересмотра методологических подходов к их исследованию, в частности, касающихся периодичности отбора проб.
Тенденции изменений бентосных сообществ
Бентосные беспозвоночные, являясь одним из основных элементов водных экосистем, участвуют в биогеохимическом круговороте многих элементов, в частности, биогенов, кальция, кремния, тяжелых металлов и трансформации органического вещества в системе «толща воды - донные отложения», что обусловливает их значительную роль в процессах самоочищения вод, регулируют газовый режим и механический состав грунтов водных объектов (Макрушин, 1984; Балушкина, 1987; Методы ..., 1989; Руководство ..., 1992; Шитиков и др., 2003).
Кольский полуостров является северным пределом распространения многих видов, однако в целом макрозообентос региона качественно богат и включает практически все систематические группы, представленные в пресных водоемах Палеарктики. Бентосная и нектобентосная фауна Мурманской области насчитывает около 500 видов беспозвоночных организмов. Основу зообентоса составляют виды европейского, европейско-сибирского и палеарктического распространения, доля космополитов невелика (11 видов) (Яковлев, 2005).
В настоящее время в сообществах фоновых водоемов отмечено сокращение биоразнообразия и роли аборигенных видов, внедрение и расселение эврибионтных видов из умеренных широт. Вселенцы заселяют преимущественно мелководные участки водоемов, бентосные сообщества в зоне профундали более стабильны, их эти изменения затрагивают в меньшей степени (Яковлев, 2005). Помимо природных факторов, эти процессы могут быть обусловлены и антропогенным воздействием. В 2011 г. в сбросном канале Кольской АЭС (оз.Имандра) обнаружена популяция крупных двустворчатых моллюсков сем. Unionidae (р. Anodonta), которые в природных водоемах региона не обитают.
Усиление темпов эвтрофикации вод, обусловленное не только антропогенным воздействием, но глобальными факторами, в том числе изменением климата в сторону потепления, приводит к росту количественных показателей донных биоценозов и повышению трофического статуса природных олиготрофных водоемов. Так, биомасса макрозообентоса оз.Малый Вудъявр, которое не подвержено прямому влиянию промышленных и хозяйственно-бытовых сточных вод, по сравнению с данными за 1930-1939 гг. увеличилась в 3 раза, трофический статус водоема повысился до а-эвтрофного (табл.1).
Антропогенное воздействие приводит к нарушениям структурной организации бентосных сообществ, которые проявляются в сокращении видового разнообразия, элиминации или снижении роли стенобионтных видов, перестройке доминантного комплекса, упрощении трофической и этологической структуры, возрастании роли беспозвоночных, образ жизни которых связан с ползанием или хождением (Яковлев, 2005). Наиболее чувствительны к ухудшению условий среды реликтовые рачки Mysis т1Ша Loven и Monoporeia ajfnis (Bousield), моллюски Pisidium conventus Clessin, Margaritifera margaritifera Ь, личинки веснянок, поденок, ручейников, хирономид п/сем. Тапуагат и Ог&ос1аШтае. На загрязненных участках образуются сообщества из эврибионтных, или устойчивых к загрязнению видов, к которым относятся личинки хирономид родов Chironomus, Procladius, олигохеты сем. ТиЬШс1<!ае, некоторые виды моллюсков родов Lymnaea, Pisidium (Яковлев, 1991).
На современном этапе макрозообентос водоемов, подверженных значительной токсической нагрузке при сопутствующем эвтрофировании (Монче-губа оз.Имандра, оз.Куэтсъярви), представлен ограниченным числом устойчивых к загрязнению вод тяжелыми металлами групп беспозвоночных. Доминируют в составе бентоса хирономиды, устойчивость которых к действию тяжелых металлов отмечена многими авторами ^аейег, 1979; Яковлев, 1998а; 2005; Ильяшук, 2002). В зонах максимального загрязнения хирономиды формируют донные биоценозы, состоящие из монокультуры р. Chironomus.
Уровень численности и биомассы таких сообществ подвержен значительным колебаниям, которые обусловлены как естественными процессами, так и антропогенными факторами. В частности, при залповых сбросах сточных вод отмечается тотальная гибель бентосных организмов (Яковлев, 2005; Моисеенко, 2009).
Снижение токсической нагрузки на водоем вследствие сокращения объемов сточных вод привело к интенсификации процессов самоочищения водных экосистем. Одним из результатов этого является реколонизация отдельными группами макрозообентоса прежних местообитаний (табл.2). Так, в Монче-губе за период с 1996 г. по настоящее время обилие амфипод Monoporea ajfns возросло в 18 раз. В 2009 г. М. affim были отмечены на расстоянии 5-6 км от источника загрязнения, что ранее не наблюдалось (Валькова, 2010).
Как было показано ранее, эвтрофирование северных водоемов не всегда приводит к деградации водных экосистем, особенно при умеренном проявлении таких процессов. На всех этапах сукцессии водоемов от олиготрофного до эвтрофного статуса водная экосистема способна адаптироваться, изменяя свою структурно-функциональную организацию и перестраивая механизмы утилизации органического вещества (Яковлев, 2005). В настоящее время макрозообентос водоемов, подверженных антропогенному эвтрофированию
(оз.Ковдор, Большой Вудъявр), характеризуется относительно высоким таксономическим разнообразием беспозвоночных - 12-14 групп ранга семейств и отрядов. Избыточное поступление биогенов приводит к возрастанию количественных показателей, увеличению доли олигохет в структуре донных биоценозов и повышению трофического статуса водоемов. Так, по данным за 2001 г. (Кашулин и др., 2009), средняя биомасса зообентоса оз.Большой Вудъявр составляла 9 г/м2, что соответствует Р-мезотрофному типу водоемов, в 2010 г. уровень биомассы донных биоценозов составлял 14 г/м2, трофический статус озера повысился до а-эвтрофного (табл.3).
Таблица 2
Параметры бентосных сообществ Монче-губы озера Имандра в разные периоды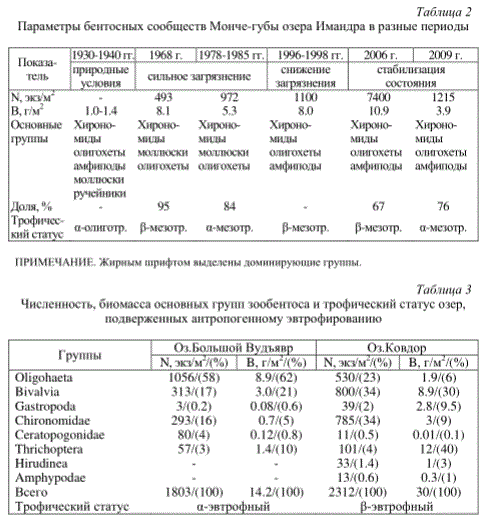
Трофический статус а-эвтрофный Р-эвтрофный
В трофической структуре макрозообентоса преобладают грунтозаглатыватели, собиратели-детритофаги и фильтраторы, группировка хищников развита слабо. Такая структура сообществ, направленная утилизацию избыточного органического вещества и биогенов, свидетельствует о преобладании детритных пищевых цепей в водоеме.
Тенденции изменений зоопланктонных сообществ
Зоопланктонное сообщество - часть экосистемы озера, тесно связанная со всеми остальными его звеньями (фито- и бактериопланктоном, рыбами, бентосом) и отражающая общее состояние водоема, служит надежным индикатором качества вод и играет важную роль в процессах самоочищения водоемов. Как любой биоценоз экосистемы характеризуется специфическим видовым составом, динамической устойчивостью, определенной, присущей ему организацией. Изменение условий существования организмов отражается на видовом составе, количественных показателях, соотношении таксономических групп, структуре популяций доминирующих видов. Зоопланктон - один из компонентов кормовой базы рыб, играет значительную роль в определении рыбопродуктивности водоема.
По данным В.А.Яшновой (1925) и О.С.Зверевой (1966), пресноводная фауна озер Крайнего Севера состоит из следующих элементов: 1) эврибионтные формы, характерные для Северного полушария; 2) арктические элементы фауны (Heterocope borealis, Daphnia arctica, Bosmina obtusirostris v. arctica); 3) морские реликты (Limnocalanus macrurus, Eurytemora lacustris); 4) элементы байкальской фауны (Eudiaptomus tibetanus). Согласно литературным данным (Рылов, 1922; Рыбы Мурманской области, 1966; Большие озера..., 1975; Решетников, 1980), зоопланктон северных озер насчитывает около 40 форм (коловратки, кладоцеры и копеподы), из которых большинство относится к эвритропным и массовым, но есть и виды, характерные только для северных и альпийских водоемов: Asplancna priodonta, Keratella cochlearis, Kellicottia longispina, Bosmina obtusirostris, Holopedium gibberum, Daphnia cristata, Thermocyclops leuckarti, Cyclops scutifer, Eudiaptomus gracilis, Heterocope appendiculata.
До последнего времени уровень жизни в водоемах Заполярья считался крайне низким, что объяснялось климатическими условиями зоны, низкой минерализацией воды, незначительным содержанием биогенных элементов и органических веществ (Баранов, 1962; Волхонская, 1966; Петровская, 1966). Например, в озерах лесотундрового ландшафта, находящихся в северо-западной части Кольского п-ова, численность зоопланктона колебалась в пределах 12.3-63 тыс. экз/м3, биомасса - 0.005-0.5 г/м3. В озерах северотаежного ландшафта развитие зоопланктона достигало более высокого уровня - численность 25.8-663.5 тыс. экз/м3, биомасса - 0.3-2.2 г/м3. В тундровых озерах биомасса составляла 0.005-0.3 г/м3 (Биологическая продуктивность ..., 1975).
Однако исследования последних лет выявили достаточно высокий уровень продуцирования планктона в этих широтах, что обусловлено влиянием как региональных (почвенно-климатических), так и локальных, действующих внутри самого ландшафта (морфометрических, гидрологических, гидрохимических, особенностей водосбора, хозяйственной деятельности человека и т.д.) факторов. Взаимодействие этих факторов формирует состав и численность планктонных организмов каждого конкретного водоема.
Современное состояние водных экосистем существенно отличается от такового, характерного периоду, предшествовавшему интенсивному индустриальному освоению региона. Высокие темпы индустриализации северных регионов в ХХ веке породили целый ряд проблем, связанных с изменениями качества окружающей среды (Кашулин и др., 2005). За последние десятилетия в период возрастания антропогенной нагрузки экосистемы, в прошлом олиготрофных водоемов претерпели существенные изменения, которые коснулись всех ее структурных компонентов.
Комплексный характер антропогенного воздействия на водоемы Кольского региона существенно затрудняет выявление наиболее информативных показателей зоопланктона как компонента экологического мониторинга и оценку влияния на сообщество отдельных факторов (токсичного загрязнения, эвтрофирования, теплового воздействия и др.). К сожалению, как уже отмечалось выше для сообществ фитопланктона, информация о зоопланктонных сообществах разнотипных малых озер региона также практически отсутствует или является отрывочной, а результаты, полученные по данным разового отбора проб методом экспресс-съемок, не могут дать представление о реальном экологическом состоянии водоема за краткосрочный период исследований. Однако по ряду озер (Имандра, Большой Вудъявр, Ковдор и др.) имеются данные многолетних мониторинговых наблюдений, позволяющие выявить ряд тенденций в развитии зоопланктонных сообществ при долговременном техногенном воздействии на водоемы.
Комбинированные эффекты токсичного загрязнения вод и эвтрофирования
выявлены в губе Монче оз.Имандра, озерах Большой Вудъявр и Ковдор.
Таксономическая структура зоопланктонного сообщества является хорошим индикатором степени загрязнения водоемов в целом или их отдельных участков. Структурные перестройки зоопланктона проявляются в снижении, а в ряде случаев исчезновении наиболее чувствительных к ухудшению экологических условий реликтов и типичных представителей фауны олиготрофных озер (L. kindtii, B. longimanus, E. gracilis, H. appendiculata) (табл.4). Их замещают и постепенно формируют состав руководящего комплекса эврибионтные мелкие виды с простыми жизненными циклами и высокой скоростью размножения (r-стратеги) - коловратки (в соотношении основных таксономических групп Rotatoria.Cladocera.Copepoda в величине общей численности и биомассы организмов они оставляют долю более 90%), что может служить хорошим признаком нарушенного нестабильного состояния экосистемы. Отмечается уменьшение средней индивидуальной массы зоопланктера сообщества (B/N). Снижение индекса видового разнообразия Шеннона по численности (Н№ит/экз) происходит не только в результате сокращения числа видов в сообществе, но и за счет усиления доминантности отдельных видов и создания монокультур из устойчивых к загрязнению форм (табл.5-7).
Доминирование устойчивых к загрязнению коловраток, присутствие в незначительных количествах «тонких» (Daphnia, Bosmina) и практическое отсутствие активных «грубых» фильтраторов (Eudiaptomus) следует считать этапом значительных изменений в функционировании зоопланктона как естественного биофильтра (BCrust/BRot<1). Это заметно снижает процессы самоочищения и ухудшает экологическое состояние водоемов. Длительное воздействие загрязняющих веществ приводит к упрощению экосистемы.
С удалением от источников интенсивного антропогенного воздействия отмечается рост ценных в кормовом отношении «тонких» фильтраторов-фитофагов кладоцер (B. obtusirostris, Daphnia spp) и активных «грубых» фильтраторов копепод (E. gracilis, H. appendiculata) (BCrust/BRot>1).
Количественные показатели зоопланктонного сообщества также проявляют определенную специфику в зависимости от степени техногенного загрязнения. Максимальные значения численности (до 346.3 тыс. экз/м3 в губе Монче оз.Имандра) и биомассы (до 3.8 г/м3 в оз.Ковдор и 4.4 г/м3 в оз.Большой Вудъявр) зоопланктона (табл.5-7) были зарегистрированы в зонах непосредственного загрязнения водоемов сточными водами медно-никелевого (ОАО «Кольская ГМК» и «Ковдорский ГОК») и апатит-нефелинового (ОАО «Апатит») производств и, согласно «шкале трофности» С.П.Китаева (1984), характерны для водоемов с повышенным типом трофности.
Таблица 4
Состав руководящего комплекса летнего зоопланктона оз.Имандра в многолетнем ряду исследований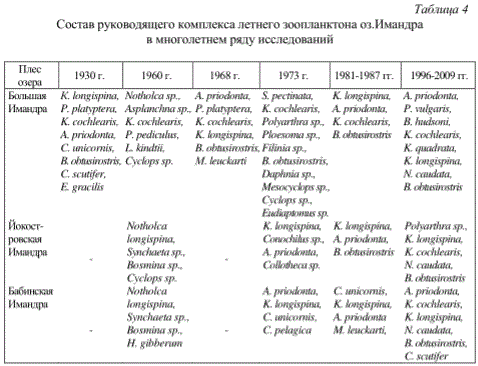
ПРИМЕЧАНИЕ. Данные за 1930 г. взяты из работ Н.И.Семеновича (1940), 1960 г. - М.В.Петровской (1966), 1968 г. - Т.В.Дольник и Г.А.Стальмаковой (1976), 1973 г. - Р.С.Деньгиной (1980), 1981-1987 гг. - Т.И.Моисеенко и В.А.Яковлева (1990), 1996-2009 гг. - Т.И.Моисеенко и др. (2009).
Таблица 5
Структурные показатели зоопланктонного сообщества в зоне распространения сточных вод медно-никелевого производства (губа Монче оз.Имандра) (гидробиологическое лето 1996, 1998, 2003, 2006 гг.)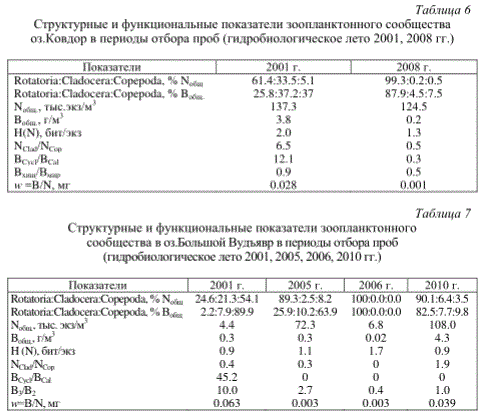
Эффекты эвтрофирования вызваны высоким содержанием биогенных элементов и органических веществ, поступающих с городскими хозяйственно-бытовыми и промышленными стоками. Согласно литературным данным (Дубровина и др., 1991), содержание больших количеств биогенных элементов и органических веществ в воде снижает токсичность тяжелых металлов и других металлов и частично стимулирует развитие зоопланктона. Как типичный признак повышения трофического статуса водоемов отмечается устойчивая тенденция замены «грубых» фильтраторов каланоид «тонкими» фильтраторами, представленными ветвистоусыми ракообразными (N^/N^>1).
Комбинированные эффекты загрязнения вод минеральной взвесью и эвтрофирования отмечены в губе Белой оз.Имандра. Ретроспективный анализ данных показал, что многолетняя динамика зоопланктона в губе Белой характеризовалась сокращением видового разнообразия, уменьшением численности и биомассы до начала 1980-х гг. и последующим увеличением количественных показателей к 1990 г. В период исследований с 1978 по 1991 гг. отмечалось массовое развитие коловраток (численность выше 70%, биомасса - 55% всего зоопланктона) (Яковлев, 1998). Вблизи дамбы отстойника АНОФ видовой состав зоопланктона был крайне обедненным, а количественные показатели низкими. К выходу в открытое озеро видовое разнообразие и количественные характеристики зоопланктона закономерно возрастали (в основном за счет коловраток). На специфическую структуру сообщества зоопланктона могло оказать влияние и обильное развитие
в воде сапрофитных, денитрифицирующих бактерий, актиномицетов и «фосфорных» бактерий, способных разлагать некоторые нерастворимые минеральные формы Робщ (Яковлев, 1998). Преобладали A. priodonta, Notholca sp., B. obtusirostris, хищные веслоногие рачки рода Acanthocyclops. Фильтраторов и седиментаторов обнаруживали только в пелагиали плеса, где концентрация минеральных тонкодисперсных частиц в воде была сравнительно низкой. Здесь наряду с A. priodonta, K. longispina, K. cochlearis и S. pectinata в пробах присутствовал рачок B. obtusirostris (Деньгина, 1980; Моисеенко, Яковлев, l990; Яковлев, 1995). С 1981 по 1990 гг. среднегодовая численность зоопланктона увеличилась почти в 9 раз (приблизительно с 9 до 81 тыс. экз/м3).
В период гидробиологического лета 1996, 1998, 2003, 2006, 2011 гг. были зарегистрированы высокие значения общей численности (491.1 и 326.5 тыс. экз/м3) и биомассы (3.4 г/м3) зоопланктона (табл.8). Доминировали коловратки, причем крупная хищная A. priodonta составляла 80-90% общей численности. Происходит замена «тонких» фильтраторов-фитофагов Bosmina и Daphnia, характерных для водоемов с повышенным уровнем трофии и не способных отфильтровывать крупные частицы взвешенных органических веществ, на хищных веслоногих циклопов (M. leuckarti, A. gigas) при общем преобладании коловраток (A. priodonta, K. cochlearis, K. quadrata, К. longispina). Известно, что коловратки, благодаря смешанному характеру питания, менее чувствительны по сравнению с кладоцерами к условиям высоких концентраций неорганической взвеси (Gliwicz, 1969; Телеш, 1996). Циклопоиды, обладая прочными хитиновыми покровами и имея хищный тип питания, также более устойчивы к воздействию токсикантов. Чувствительные к загрязнению активные «грубые» фильтраторы-фитофаги каланоиды (E. gracilis), изымающие из толщи воды крупные частицы взвешенных органических веществ, были отмечены единично, что свидетельствует о снижении биофильтрационной активности зоопланктона в данном районе озера.
Таблица 8
Структурные показатели зоопланктонного сообщества в зоне распространения сточных вод апатитонефелиновых обогатительных фабрик (губа 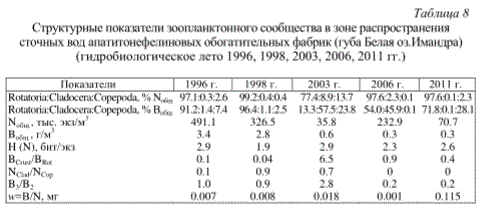
В зоне влияния подогретьх вод Кольской АЭС (губа Молочная оз.Имандра) отмечается увеличение доли ветвистоусых и веслоногих ракообразных. Отличие теплового воздействия от техногенного загрязнения и эвтрофирования водоемов заключается в том, что в первом случае в водоемы поступает тепло (энергия), а в остальных - вещество (Мордухай-Болтовской, 1975). Тепло не аккумулируется в водоемах и не передается по трофическим цепям. Существенно отличается и период последействия. После прекращения сброса теплых вод за время, необходимое на перераспределение субстратов, восстанавливаются даже полностью уничтоженные
высокими температурами биоценозы. После прекращения сброса загрязняющих веществ необходимо длительное время проводить специальные мероприятия по очистке водоемов, и лишь потом становится возможным восстановление разрушенных биоценозов (Кошелев, 1983; Здановски, 1991). Также рядом исследователей было доказано, что в водоемах Кольского п-ова термальное воздействие отработанных вод электростанций приводит к изменениям в сообществах, в определенных пределах сходным с влиянием эвтрофирования. Повышение температуры способствует ускорению процессов роста и развития гидробионтов: происходит более раннее созревание и размножение многих организмов, удлиняется их вегетационный период (Niva, 1973; Gallup, Hickman, 1975; Крючков и др., 1985; Моисеенко, 1997).
Подогреваемая акватория оз. Имандра (губа Молочная), несмотря на наметившийся процесс эвтрофирования, сохраняет в основном черты олиготрофного водоема (численность 7.7-87.0 экз/м3, биомасса 0.1-1.1 г/м3) (табл.9), что подтверждает выводы ряда исследователей о том, что влияние умеренно подогретых сбросных вод электростанции на животный и растительный мир субарктического водоема имеет в целом положительный фактор.
Таблица 9
Структурные показатели зоопланктонного сообщества в зоне влияния подогретых вод Кольской АЭС (губа Молочная оз.Имандра) (гидробиологическое лето 1996, 1998, 2003, 2006, 2011 гг.)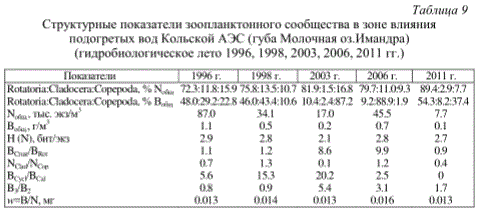
В качестве отрицательного эффекта следует отметить гибель и травмирование части крупных, имеющих выросты ракообразных (фитофагов Daphnia, Bosmina, факультативного хищника Cyclops и облигатных хищников Leptodora, Bythotrephes) при прохождении через охладительную систему Кольской АЭС, а также влияние сложной гидродинамической ситуации, обусловленной высокой степенью перемешивания и проточностью водных масс в устье сбросного канала и подогреваемой зоне озера (табл.4). Негативные последствия теплового воздействия на сообщества гидробионтов в условиях Субарктики могут выражаться и в снижении самоочищающей способности экосистемы озера. Последнее особенно опасно, так как вокруг оз.Имандра расположены крупные добывающие и перерабатывающие комплексы, а также населенные пункты, сбрасывающие в озеро частично очищенные и неочищенные стоки.
Особого внимания в настоящее время заслуживает проблема «цветения» воды в оз.Имандра, связанная с массовым развитием цианобактерий вследствие процессов эвтрофирования. Это явление само по себе является мощным стрессором для экосистемы и создает множество проблем при рекреационном, хозяйственном и питьевом использовании водоема. При цветении воды происходит резкое снижение концентрации растворенного кислорода, что негативно сказывается на жизнедеятельности гидробионтов, вплоть до их гибели.
Зоопланктонное сообщество «цветущего» водоема также претерпевает структурно'-функциональные изменения. Ряд исследователей отмечают повышенное содержание мертвых особей и различные патологии на видовом и популяционном уровне (Семенова, 2009), аномалии, визуально зафиксированные в процессе обработки проб (бледная окраска особей, раскрытые створки и опухолеобразные наросты у Cladocera, распавшиеся колонии у представителей рода Conochilus) (Андроникова, 2007). Под влиянием токсинов синезеленых водорослей увеличивается образование эфиппиев у ветвистоусых (Breitholtz et al., 2001). Также было выяснено, что токсическое действие может быть одной из причин, вызывающей абортирование молоди и развитие аномалий у ракообразных (Carmichael, 2001). Периоды гиперцветения являются неблагоприятными для зоопланктона, и различные показатели зоопланктонного сообщества могут рассматриваться как индикаторы состояния водной среды в период интенсивного развития цианобактерий.
По мнению ряда авторов (Яковлев, 1991; Кашулин и др., 2005; Моисеенко и др., 2009), экосистемы переходят к новой модификации, отличной от их природной структуры, и не происходит возвращение их к природному состоянию.
Тенденции изменений рыбной части сообщества субарктических водоемов
При всем многообразии и многочисленности внутренних водоемов высокоширотной области Северо-Запада России, в отличие от озер центральных и южных областей, видовое разнообразие ихтиоценозов здесь, как правило, значительно ниже. Ихтиофауна водоемов Северной Фенноскандии характеризуется доминированием в составе сообществ представителей стенобионтных лососевых и сиговых рыб. Достаточно широко распространены окуневые, щуковые, а в бассейнах крупных речных систем - также и колюшковые. По различным оценкам, в состав ихтиофауны данного региона могут входить 28 видов и подвидов пресноводных, проходных, полупроходных, морских и солоноватоводных рыб (Берг, Правдин, 1948; Галкин и др., 1966; Сурков, 1966; Лукин, 1998; Муравейко и др., 2000; Атлас..., 2003; Калюжин, 2003; Карамушко, Берестовский, 2005). Однако в озерных экосистемах ихтиофауна чаще представлена 5-9 основными видами.
В настоящее время структура рыбной части сообщества субарктических водоемов претерпевает существенные изменения. Это связано радикальными изменениями условий обитания рыб под воздействием глобальных и региональных факторов. В большинстве случаев в природных водоемах мы имеем дело с сублетальными продолжительными воздействиями поллютантов. Отрицательные эффекты могут проявляться на различных уровнях биологической организации: от субклеточного до популяционного и сообществ. Для понимания последствий воздействия токсикантов на состояние популяции и прогноза ее существования необходимо изучать эффекты веществ-загрязнителей на различных уровнях биологической организации. Большинство популяционных показателей не всегда может однозначно раскрыть причины изменений состояния популяции в силу неспецифичности их возможных ответов на различные типы стрессов. Особенно сложно выделить влияние загрязнений на популяционные характеристики рыб при их хроническом сублетальном и/или многофакторном антропогенном воздействии. Изменения популяционных показателей, если не известен их источник, не могут быть однозначно интерпретированы без комплексной информации о состоянии организма, что делает необходимым проведение соответствующих исследований.
Особое место среди показателей состояния организма занимает содержание веществ-загрязнителей в его органах и тканях. Содержание веществ-загрязнителей в организме гидробионотов, отражая общую нагрузку, определяется сложными процессами поглощения, перераспределения внутри организма, детоксикации и выведения из организма. В настоящее время, помимо локальных источников, увеличение нагрузки загрязняющих веществ на водоемы может происходить за счет процессов глобального загрязнения атмосферы и трансграничного переноса. Примерами такого рода процессов являются распространение свинца и ртути. Нами было показано, что содержание ртути в донных отложениях озер и тканях рыб Мурманской области и приграничного района Финляндии и Норвегии за последнее десятилетие характеризуется постоянным ростом и этот процесс не связан с деятельностью металлургических комбинатов региона (рис.6) (State ., 2007).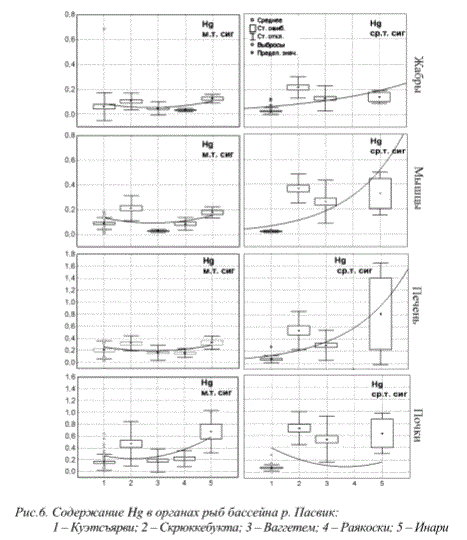
Рис. 6. Содержание Hg в органах рыб бассейна р. Пасвик:
1 - Куэтсъярви; 2 - Скрюккебукта; 3 - Ваггетем; 4 - Раякоски; 5 - Инари
Несмотря на это, по-прежнему приоритетным элементом-загрязнителем региона остается никель. Его концентрация, в отличие от других элементов, в почках и жабрах рыб хорошо коррелирует с антропогенной нагрузкой на водоемы (рис.7).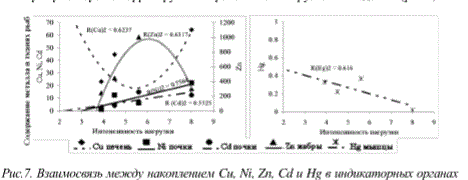
Рис. 7. Взаимосвязь между накоплением Си, №, 7п, Cd и Щ в индикаторных органах сига и уровнем техногенной нагрузки на исследованные водоемы (натуральный логарифм концентрации М в поверхностном слое донных отложений)
Никель способен влиять на содержание в тканях многих других элементов. Можно полагать, что именно никель является основным патогенным фактором в исследованных водоемах. Уровень 5-7 мкг/гсухв-ва в почке, вероятно, является критическим для сигов изученных водоемов, и его превышение вызывает ряд серьезных патологических процессов в организмах рыб.
В условиях продолжительного аэротехногенного загрязнения вод тяжелыми металлами, его региональный характер подчеркивает и тот факт, что уровни накопления ряда металлов (например Си, 2п) в организмах рыб в регионально-фоновой зоне зачастую имеют сопоставимые, а иногда и более высокие показатели, чем у рыб вблизи промышленных центров (Куэтсъярви) (рис.8).
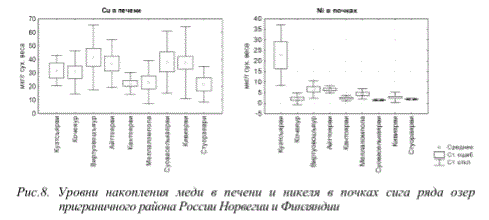
Рис.8. Уровни накопления меди в печени и никеля в почках сига ряда озер приграничного района России Норвегии и Финляндии
Действие загрязняющих веществ вызывает у рыб стресс, при этом в их организме происходят внутренние изменения, которые могут быть вредными или адаптивными. Стрессоры окружающей среды, в т.ч. и антропогенные, вызывают изменения в клеточной функции, что ведет к изменениям физиологии систем органов в индивидуальном организме. Зачастую сублетальный эффект первоначально определяется биохимическими процессами, и большинство
проявляющихся на организменном уровне биологических ответов базируется на взаимодействии между токсикантами и структурными или/и функциональными компонентами клетки. Такие первичные взаимодействия индуцируют последующие структурные и функциональные изменения на более высоких уровнях организации, проявляясь в нарушении жизненно важных функций, таких как нервные и мышечные, дыхательные, обменные, иммунные, осморегуляторные и гормонорегуляторные. Эти эффекты ведут к необратимым нарушениям различных процессов, протекающих в организме, и его функций - поведение, рост, репродукция, выживание и т.д., что является причиной значительных изменений на уровне популяций и сообществ в целом (изменение структуры и численности популяций, их исчезновение, нарушение видового разнообразия и т.д.), так как состояние популяции рыб является в конечном счете результатом ответов отдельных организмов на изменение окружающей среды.
Тяжесть и характер патологического процесса у рыб, вызванного различными токсическими веществами, можно установить на основе клинической и патолого-анатомической диагностики (Лукьяненко, 1987). Для подтверждения причин стресса у популяций сигов и оценки состояния организмов рыб были проведены патолого-морфологические исследования, включающие как клинический осмотр рыб, так и их патолого-анатомический анализ. В полевых условиях этот метод является высоко информативным, не требующим дорогостоящего оборудования, и позволяет выявить отклонения функционирования жизненно важных органов (Канаев, 1985; Аршаница, Лесников, 1987; Чинарева, 1988; Моисеенко, Яковлев, 1990; Аршаница, 1991).
Состояние организмов рыб в исследованных нами водоемах свидетельствовало о сильном стрессе. Наблюдаемые патологии характерны для водоемов, загрязняемых тяжелыми металлами, и свидетельствуют о токсичности окружающей среды. Практически во всех водоемах региона в организмах рыб происходят патологические изменения, обусловленные сублетальной токсичностью среды обитания и в целом снижающие их жизнеспособность. Хроническое воздействие загрязняющих веществ вызывает в организмах рыб нарушение функций таких жизненно важных органов, как жабры, печень, почки, гонады. Частота встречаемости патологий у рыб и их интенсивность имеет выраженную зависимость от величины техногенной нагрузки на водоем (рис. 9).
И патологии жабр ■ патологии гонад
□ патологии почек □ нефрокальцитоз
□ патологии печени
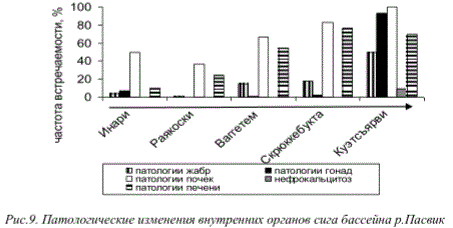
Рис.9. Патологические изменения внутренних органов сига бассейна р.Пасвик
Многолетние наблюдения за состоянием организмов рыб в районах непосредственного влияния предприятий цветной металлургии свидетельствуют о том, что частота встречаемости патологий органов и тканей не снижается. Так, для водоемов бассейна река Пасвик - водохранилищ Ваггетем и Скрюккебукта, распложенных на удалении около 40 и 16 км соответственно от предприятия «Печенганикель», в настоящее время сохраняется весьма высокий процент патологий внутренних органов сига. Необходимо отметить, что на протяжении нескольких лет наблюдений, отмечена тенденция к росту частот встречаемости видоизменений репродуктивной системы рыб и печени, несмотря на проводимые в этот период мероприятия по снижению предприятием выбросов вредных веществ (рис. 10)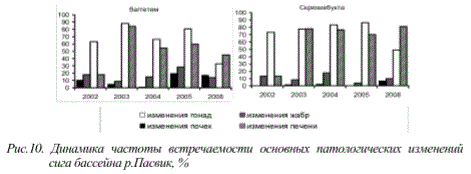
Рис. 10. Динамика частоты встречаемости основных патологических изменений
сига бассейна р.Пасвик, %
И если раньше патологические изменения внутренних органов рыб были характерны лишь для водоемов, расположенных в так называемых «импактных» зонах крупных горноперерабатывающих и металлургических предприятий, то в настоящее время в той или иной степени они наблюдаются практически по всей территории Мурманской области и частота их встречаемости возрастает.
Так, наблюдения на оз.Кочеявр (водосбор р.Лотта, приграничный района России и Финляндии), проводимые с начала 90-х годов ХХ века, показали, что если начальный период наблюдений у сигов отмечались главным образом изменения жабр, печени и почек, то уже в последующие 1991-1992 гг. регистрировался рост патологий печени в виде изменения ее окраски и появления мозаичной структуры (с 83 до 100%). Около 27% особей имели патологии гонад в виде перетяжек и перекрученной структуры (самцы), а также студенистости, слабой пигментации икринок (самки) и ассиметричности парных гонад. Частота встречаемости заболевания почек с 1991 г. (8%) также резко увеличивалась в 1992 г. и охватывала же около трети особей (Кашулин, 1994; Кашулин и др., 1999). За последнее десятилетие произошло выравнивание частоты встречаемости основных патологий. Тем не менее, основной процент патологических изменений сигов озера по-прежнему приходится на жизненно важные органы - печень и почки.
Характер патологий рыб, отмечаемых в интенсивно загрязняемых водоемах, имеет сходные черты с изменениями органов и тканей представителей ихтиофауны так называемых фоновых районов региона - центральной части Кольского полуострова, расположенных в зоне регионального фонового содержания основных поллютантов медно-никелевого производства. При этом частота встречаемости патологических трансформаций внутренних органов рыб в водоемах северной лесной зоны Финляндии и Мурманской области, находящихся на значительном удалении от источников аэротехногенного загрязнения, в настоящее время достаточно высока (рис. 11).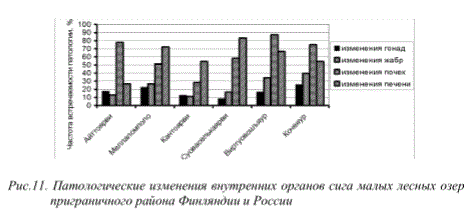
Рис.11. Патологические изменения внутренних органов сига малых лесных озер приграничного района Финляндии и России
У сигов среднего течения бассейна р.Поной также была отмечена высокая частота встречаемости патологии жабр, печени и почек (рис.12). Однако интенсивность изменений органов здесь имеет начальный характер, что, вероятно, также обусловлено долговременными процессами аэротехногенного загрязнения водоемов. Очевидно, что в настоящее время специфика и интенсивность патологических изменений рыб исследованных водоемов, испытывающих разноуровневую аэротехногенную нагрузку, приобретают аналогичный характер.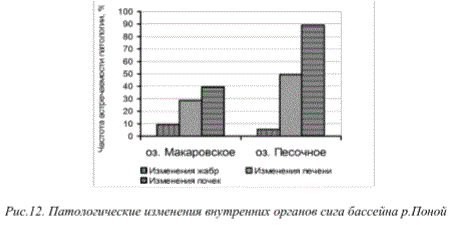
Рис.12. Патологические изменения внутренних органов сига бассейна р.Поной
Во взаимодействиях с окружающей средой сложность и разнообразие популяционной структуры определяют ее устойчивость и жизнеспособность. Популяция как единая биологическая система структурирована пространственно и функционально, что позволяет ей эффективно использовать имеющиеся в ее распоряжении ресурсы - как энергетические, так и пространственные. Она функционирует как целое благодаря взаимодействию организационных элементов (Алимов, 1989). Разнокачественность особей в составе внутрипопуляционных группировок имеет огромное значение в формировании адаптационного ответа популяции на меняющуюся экологическую обстановку (Шилов, 1985; Жизнеспособность популяций ..., 1989; Кудерский, 1991). Разные возрастные категории
животных и различные внутривидовые формы, обитающие в одном водоеме, имеют и генетические особенности. Упрощение структуры популяции снижает возможности популяции в воспроизводстве и в освоении различных ниш, эффективность использования ею ресурсов экосистемы и генотипическое разнообразие популяций.
Для популяций рыб, обитающих в исследованных нами водоемах, характерно крайнее упрощение их структуры. Они представлены небольшим числом возрастных групп и минимальным числом нерестящихся генераций (рис. 13). Наблюдается сокращение продолжительности жизни, преобладание рыб младших возрастных групп, снижение темпов роста и уменьшение средних размеров, раннее половое созревание, наступление его при экстремально малых для вида размерах или/и блокировка процессов созревания при увеличенных темпах роста, растянутый период 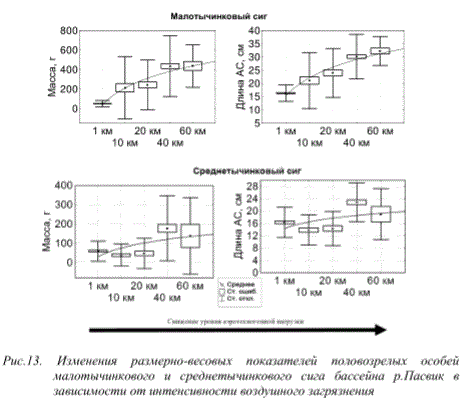
Рис.13. Изменения размерно-весовых показателей половозрелых особей малотычинкового и среднетычинкового сига бассейна р.Пасвик в зависимости от интенсивности воздушного загрязнения
Анализ ряда популяционных показателей рыб, обитающих в водоемах импактных зон, испытывающих значительную нагрузку техногенных загрязнителей, показывает, что стресс, обусловленный токсичностью окружающей среды, вызывает сокращение продолжительности жизни рыб и снижение темпов роста. Происходит резкое уменьшение числа возрастных групп и генераций, способных к нересту (фактически до одной). Имеет место резкое омоложение начала полового созревания рыб и наступление его при экстремально малых для вида размерах. Так, сиги оз.Куэтсъярви нерестятся на втором году жизни при длине тела 7-9 см. При этом происходит резкое снижение абсолютной плодовитости. Это дает право говорить, что
имеет место смещение динамики численности популяций к короткому моноциклу. Наблюдается нарушение соотношения соматического и генеративного обменов в пользу преобладания последнего. Как следствие этого - ускоренное созревание (в более раннем возрасте и при меньших размерах) и образование карликовых форм.
Известно, что обычно созревание рыб наступает при достижении определенных размеров и быстрорастущие особи созревают в более раннем возрасте, а тугорослые - позднее (Васнецов, 1953а, б). Для северных регионов характерно более позднее созревание рыб, которое объясняется более медленными темпами накопления "резервных белков" и достижением определенного уровня жирности, необходимых для обеспечения нормального созревания половых продуктов (Решетников и др., 1989). В первой половине прошлого века для сигов нормой считалось наступление первого полового созревания в возрасте 5+-6+ лет, при продолжительности жизни рыб до 23-27 лет. Для нереста рыбам необходим определенный запас энергии и веществ. В то же время при меняющихся условиях обитания зависимость между темпами роста и возрастом наступления половой зрелости может изменяться, что связывают с изменениями характера обменных процессов (Лапин и др., 1985). Ускорение энергетического обмена, обусловленное изменением среды обитания, ведет к увеличению скорости роста и более раннему созреванию сиговых, но и к снижению предельных размеров (Канеп, 1981). Наблюдаемое нами раннее созревание сигов, вероятно, является ответной реакцией популяции на сокращение продолжительности жизни в результате повышения токсичности среды. Оно происходит на фоне снижения темпов роста рыб и ухудшения состояния их организмов.
Изменение стратегии жизненного цикла, направленное на сохранение жизнеспособности популяции в целом и позволяющее поддерживать достаточно высокую численность, служит ответом популяции на субтоксичность окружающей среды. Процессы детоксикации тяжелых металлов и раннее половое созревание требуют повышенных энергетических затрат, что обеспечивается достаточностью пищевых ресурсов. В то же время в уловах могут присутствовать крупные, хорошо упитанные особи, большая часть которых не участвует в нересте. Т.И.Моисеенко (1997) на основании анализа популяций рыб больших водоемов Кольского полуострова, испытывающих мощное техногенное загрязнение, выделяет два основных типа ответов, обусловленных увеличением расходов энергии на детоксикацию:
• замедление темпа роста при интенсивном питании и повышенном жиронакоплении, отсрочка созревания и более частые пропуски нерестового сезона;
• переход на короткий цикл воспроизводства и стремление к моноциклии.
При первом типе реакций энергетические затраты на поддержание
гомеостаза популяции в субтоксичных условиях среды «оплачиваются» отсрочкой созревания и рассматриваются как переходное состояние популяции, запускающее механизмы эволюционной предадаптации популяций к стрессу. С усилением токсичной нагрузки (повышающей элиминацию особей старших возрастных групп и необходимость в увеличении «энергетических плат» за детоксикацию) долгожительство и позднее созревание становятся экологически невыгодными. Популяция стремится к новому стабильному состоянию, выражающемуся в переходе на укороченный жизненный цикл. Адаптивную ценность приобретает способность популяции сокращать энергоемкие функции: долгожительство, соматический рост, высокую частоту нереста, что проявляется в «сжимании» структурных рядов. Ресурсы, которыми располагает организм в субтоксичных условиях, ограничены большими затратами на процессы детоксикации. Как известно,
репродуктивные траты также являются чрезвычайно энергоемкими и текущее размножение часто подавляет выживаемость, рост или будущее размножение (Бигон и др., 1989). Таким образом, ограничение ресурсов и снижение под воздействием тяжелых металлов эффективности многих функций организма обусловливают образование карликовых короткоживущих форм, что можно рассматривать как компромисс в расходовании доступной энергии.
«Переход» на короткий цикл развития в условиях достаточного обеспечения пищи позволяет рыбам поддерживать высокую численность популяций даже в самых загрязненных водоемах, если процесс техногенного загрязнения сопровождается процессами эвтрофикации (оз.Куэтсъярви, губа Белая оз.Имандра и др.). Однако присутствие в популяциях минимального числа нерестящихся генераций (одной-двух) в условиях усиления техногенного стресса и/или появления новых стрессовых факторов делает проблематичным длительное существование этих популяций. Для рыб исследованных водоемов характерно сокращение численности старших возрастных групп. Кроме того, наблюдается большое количество рыб, пропускающих нерест. Это позволяет сделать предположение, что в основном воспроизводство идет за счет впервые нерестующих рыб и находится в критическом состоянии. Ухудшение качества среды или нормальные колебания природных факторов могут привести к катастрофическим последствиям. Вселение новых видов рыб, более устойчивых к изменяющимся условиям обитания, создает дополнительное напряжение в пищевых отношениях в рыбной части сообщества, что в условиях субтоксичной нагрузки является мощным дополнительным стрессом для популяций аборигенных видов.
Стратегия, характеризующаяся отсрочкой созревания с возможной реализацией размножения в более поздние сроки, оказывается малоэффективной в субтоксичных условиях среды. Она наиболее распространена в популяциях, обитающих в водоемах, испытывающих относительно небольшой уровень техногенной нагрузки (Кочеявр, Ваггетем), что можно расценивать как первичную реакцию популяций на загрязнение. Особи, стремящиеся к моноциклии, имеют явные преимущества, так как они в состоянии реализовать репродуктивный потенциал в течение жизненного цикла и сохранить жизнеспособность популяции, даже в условиях высокой нагрузки (Куэтсъярви). Однако эффективность этой стратегии будет определяться в значительной мере количеством доступной энергии и реализуется только в водоемах с высоким уровнем трофности.
Наряду с указанными выше особенностями изменений стратегий жизненного цикла рыб водоемов Мурманской области в изменяющихся условиях окружающей среды, в последние годы отмечается совершенно новый феномен «гигантизма» короткоцикловых видов рыб. Известно, что продолжительность жизни таких видов относительно невелика, а их размерно-весовые характеристики, как правило, не превышают определенных величин и могут изменятся лишь в зависимости от географического расположения водоемов. Для субарктических олиготрофных водоемов обычно эти показатели меньше, чем для крупных бассейнов внутренних вод умеренной зоны и южных частей ареала распространения. Поэтому явления экстремально высоких для вида размерно-весовых показателей короткоцикловых видов рыб Мурманской области могут свидетельствовать о значительных трансформациях экосистем, при этом они не всегда регистрируются в благополучных с экологической точки зрения водоемах. К числу указанных видов можно отнести ряпушку оз.Ковдор (бассейн оз.Имандра), оз.Кахозеро (бассейн р.Кола), а также ерша оз.Колозеро (бассейн р.Кола) и Пермусозеро (бассейн оз.Имандра) (рис. 14).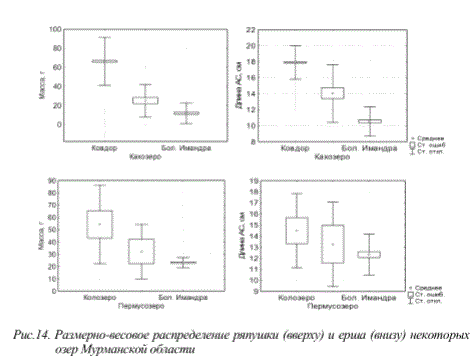
Рис.14. Размерно-весовое распределение ряпушки (вверху) и ерша (внизу) некоторых
озер Мурманской области
Подобные явления обусловлены тем, что в определенных условиях для указанных видов реализация воспроизводства и роста оказывается более успешной по сравнению с другими рыбами. При отсутствии достаточного количества хищников и большей устойчивости к изменившимся условиям обитания данные виды выигрывают конкурентные пищевые взаимоотношения с видами, занимающими аналогичные с ними экологические ниши, но чувствительными к качеству среды.
В последние годы в популяциях сигов, несмотря на снижение уровня техногенной нагрузки, сохранилась негативная тенденция снижения продолжительности жизни рыб. Признаки деградации популяций рыб (омоложение популяции за счет снижения продолжительности жизни, снижение темпов роста, переход на короткий жизненный цикл, образование карликовых форм, неравномерность ее возрастной структуры, раннее половое созревание особей, отсрочка созревания и блокировка полового развития у быстрорастущих особей, интенсивное развитие патологий внутренних органов и т.д.) прослеживаются как вблизи промышленных предприятий, так и в значительно удаленных районах (рис. 15, 16).
Например, в популяциях сига, обитающих в озерах бассейна р.Поной, несмотря на их значительное удаление от промышленных центров региона, нами было также отмечено небольшое число возрастных групп в целом и в т.ч. групп, принимающих участие в нересте. При этом среди половозрелой части популяции рыб данного вида отмечен достаточно высокий процент рыб, пропускающих нерест (рис.17). Отсутствие в выборках рыб младших возрастных групп указанных озер может быть связано как с обособлением территорий нагула и нереста в пределах озерно-речных систем, так и с возможными нарушениями воспроизводства популяции. Таким образом, внутрипопуляционные изменения рыбной части сообществ, характерные для интенсивно загрязняемых водоемов, отмечаются практически повсеместно на всей территории Мурманской области.
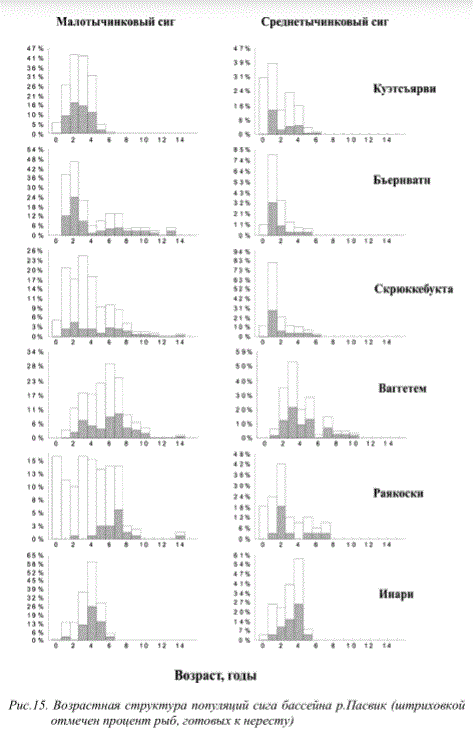
Рис.15. Возрастная структура популяций сига бассейна р.Пасвик (штриховкой отмечен процент рыб, готовых к нересту)
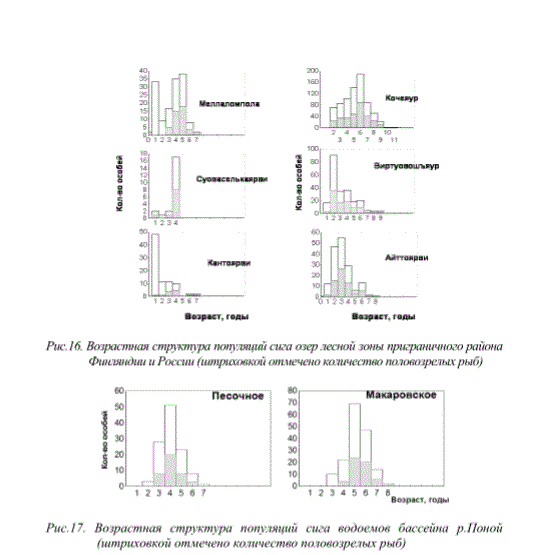
Рис.16. Возрастная структура популяций сига озер лесной зоны приграничного района Финляндии и России (штриховкой отмечено количество половозрелых рыб)
Рис.17. Возрастная структура популяций сига водоемов бассейна р.Поной (штриховкой отмечено количество половозрелых рыб)
Хорошо известно, что структурная и функциональная организация экосистем, обеспечивающая их стабильность во времени и устойчивость к изменениям внешней среды, в том числе и под воздействием антропогенных факторов, определяется биологическим разнообразием (Алимов, 1997). Поэтому изменение видовой структуры экосистем может служить показателем изменений условий обитания (Karr, Dudley, 1981; Karr, 1987; Fausch et al., 1990). Однако использование видового разнообразия для оценки деградации водоема имеет определенные сложности, связанные в первую очередь с неравномерностью распределения видов в нормальных условиях, обусловленной различными абиотическими, биотическими, историческими факторами (гидрологические, температурные условия, уровень трофности водоема, сезонность распределения, паразитарные инвазии, инфекции, историческое распространение вида и т.д.). Особенно проблематично использование этого методического подхода к водоемам высоких широт, где видовое разнообразие ихтиофауны невелико и обилие видов заменяется обилием внутривидовых форм. Сравнение видового состава затрудняется также ландшафтным и морфометрическим
разнообразием водоемов этого региона, зачастую определяющим видовой состав обитающих в них организмов. Видовой состав вблизи расположенных водоемов может резко различаться. Поэтому при изучении изменений видового состава ихтиофауны субарктических и арктических водоемов предпочтительнее преимущественно опираться на исторические данные (доиндустриального периода).
Несмотря на пиковые нагрузки загрязнений во второй половине прошлого века, длительное время структура рыбной части сообществ даже наиболее техногенно-трансформированных водоемов в целом оставалась малоизмененной. Как правило, отмечалось снижение доли лососевых видов и доминирование сиговых. Однако в конце 90-х годов прошлого и в начале нынешнего столетий во многих водоемах региона в условиях региональных изменений климата, наметились тенденции изменения в структуре сообщества рыбного населения, характерные для интенсивно эвтрофируемых водных объектов.
Изменение видового состава рыб и/или его структуры происходит несколькими путями: а) вселение новых видов; б) резкая смена доминирующих аборигенных видов; г) совместное протекание первых двух процессов. К негативным факторам, определяющим современный статус ихтиофауны водоемов Северной Фенноскандии, следует отнести загрязнение вод, изменения гидрологических условий водоемов вследствие зарегулирования стока крупных рек, интенсивное рыболовство (как промышленное, так и «любительское»), целенаправленное или стихийное вселение новых видов. Особо следует выделить резкое ускорение процессов эвтрофикации водоемов, чему может способствовать потепление климата. В целом эвтрофикация водоемов озерного типа относится к естественным процессам «старения», ввиду избыточного количества биогенных веществ и чрезмерной продукции органического вещества. Однако в условиях антропогенного эвтрофирования скорости изменения экосистем значительно возрастают. При этом трофический статус водоема также увеличивается, отмечается сокращение биологического разнообразия, общее снижение качества вод.
Считается, что развитие рыбной части сообществ водоемов в естественных условиях характеризуется последовательной сменой доминирующих видов, связанной с этапами природного «старения» водоемов. Естественная скорость сукцессии может составлять сотни лет. Смена рыбной части сообщества происходит чаще всего в такой последовательности: лососевые^-сиговые^-корюшковые^-окуневые^-карповые. В условиях интенсивного антропогенного пресса на водные экосистемы субарктических водоемов изменение их трофического статуса идет гораздо более стремительными темпами. В структуре зоопланктона и рыбного населения происходит замещение крупных и долгоживущих форм на мелкие и раносозревающие. Ценные промысловые рыбы с длинным жизненным циклом заменяются «сорными» рыбами с высоким уровнем воспроизводства и высоким приростом продукции. (Антропогенное ..., 1976; Решетников и др., 1982; Жаков, 1984).
Изменения трофического статуса крупных озер европейской части России хорошо известны (Антропогенное ..., 1976; Болотова и др., 1996). В то же время считалось, что районы Крайнего Севера (Мурманская область) практически не подвержены развитию процессов эвтрофирования водоемов. Тем не менее, тенденции изменения структуры сообщества оз.Имандра (снижение численности лососевых и доминирование короткоцикловых сиговых видов) были отмечены и ранее. Такая ситуация оставалась практически неизменной до конца прошлого столетия.
В конце ХХ - начале XXI веков во многих водоемах региона отмечаются резкие изменения в продукционных процессах и структуре их сообществ, включая рыбную часть. Так, в последние десятилетия в оз.Имандра получила массовое распространение корюшка Osmerus ереНапш, став доминирующим видом. Она практически полностью вытеснила ряпушку и снижает эффективность воспроизводства остальных видов, массово уничтожая молодь, создает повышенную пищевую конкуренцию. Нерестовая стратегия корюшки, идущей на нерест в реки, оказалась эффективней местных весенне-нерестящихся видов (щука, окунь, язь) которые вследствие зимне-весенней сработки воды Нивскими ГЭС практически полностью лишаются нерестилищ в озере. Короткий жизненный цикл, отсутствие пресса хищников, малоэффективное промысловое изъятие, успешное воспроизводство делают корюшку доминирующим видом оз.Имандра. Резко возрастает и численность ерша Оутпосерка1ш сетиш. В ряде районов озера (северные районы Большой Имандры) эти два вида полностью доминируют в структуре ихтиофауны. Воспроизводство остальных видов малоэффективно, и пополнение их популяций идет в основном за счет мигрантов из придаточных озерно-речных систем (полупроходная форма сига, кумжа).
Последние данные о состоянии фауны рыб южной части плеса Йокостровская Имандра, а также плеса Бабинская Имандра показали, что, наряду с корюшкой, окунь здесь сохраняет высокую численность. Это связано с достаточным количеством благоприятных для его нереста районов в многочисленных придаточных мелководных озерно-речных системах. В то же время для щуки, по-видимому, процессы воспроизводства, ввиду флуктуаций уровневого режима вод, не столь эффективны. Наряду с изменениями гидрологического режима водоема каскадом ГЭС, значительный вклад в формирование современного облика фауны рыб этого района озера вносит термофикация вод Кольской АЭС. Ихтиофауна указанных районов оз.Имандра, считавшихся ранее крупнейшими нерестовыми угодьями арктического гольца и сига, в настоящее время претерпела значительные изменения. Так, арктический голец на данный момент в плесе Бабинская Имандра отмечается единично. Также значительно сократилась численность сига. Большее распространение в данном районе акватории озера, наряду с корюшкой, приобретает окунь. Корюшка, в свою очередь, находясь в условиях благоприятного кормового климата, при отсутствии серьезного пресса хищников, за счет высоких темпов роста достигает значительных размеров, и сама ведет хищный образ жизни. Основным кормовым компонентом ее при этом становится ряпушка. Численность последней в исследованном районе становится крайне низкой. В зоне подогретых вод уже несколько десятилетий поддерживается самовоспроизводящаяся популяция карпа Сурппш сагрО сагрО, стихийно заселенного в конце 70-х годов прошлого века (рис. 18).
Середина XX века ___________2011 г.__________
Рис. 18. Изменения в структуре рыбной части сообщества плеса Бабинская Имандра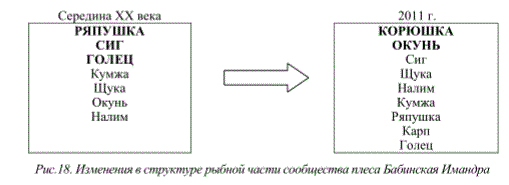
Европейская корюшка Osmerus ерег1апт, ранее обитающая в ограниченном числе водоемов бассейна Белого моря, в настоящее время освоила большее количество водоемов, успешно конкурируя с аборигенными видами рыб. В период с 1979 по 1985 гг. была выполнена интродукция онежской корюшки (в виде личинок) в Верхнетуломское водохранилище (бассейн Баренцева моря), откуда в настоящее время она также распространилась и по всей протяженности р.Кола.
Недавние исследования по оценке состояния ихтиоценозов некоторых крупных водоемов центральной части Мурманской области, относящихся к бассейнам Белого и Баренцева морей, выявили значительные перестройки в сообществах рыб, характер которых свидетельствует об изменениях в трофической структуре экосистем. Было установлено, что в исследованных водоемах доминирующими видами являются малоценные, с промысловой точки зрения, виды (рис.3). Так, в структуре ранее считавшихся лососево-сиговыми водоемами бассейнов рек Нива (Имандра, Пермусозеро) и Кола (Колозеро, Кахозеро) доминирующими видами в настоящее время являются обыкновенный ерш Оутпосерка1ш сетит, европейская корюшка Osmerus ерег1апи и европейская ряпушка Coregonus а1Ьи1а (рис.19).
Рис.19. Современная структура рыбной части сообществ некоторых водоемов Мурманской области
На сложившуюся структуру рыбного населения указанных водоемов, несомненно, оказывает влияние весь комплекс антропогенных факторов, характерных для центральных промышленных районов Мурманской области, включая неконтролируемый «любительский» лов (в основном лососевых и сиговых видов), снижение эффективности воспроизводства рыбных запасов. Это привело к снижению доли ценных промысловых пород рыб, а также хищников и, как следствие, к смене доминирующих видов в рыбной части сообщества, которыми стали такие виды, как
обыкновенный ерш и европейская корюшка. В отсутствие крупных пелагических хищников молодь ряпушки является кормовым объектом корюшки и крупного ерша оз.Колозеро, где он достигает рекордных для вида размерно-весовых показателей. Аналогичные процессы характерны для Кахозера и Пермусозера.
В стрессовых для аборигенных видов условиях вселение новых видов рыб, обладающих широкой экологической валентностью, приводит к радикальным изменениям структуры ихтиоценозов. Так, вселение ряпушки внесло значительные изменения в структуру рыбной части сообщества водоемов системы р.Пасвик. Европейская ряпушка Coregonus albula, акклиматизированная в финском оз.Инари, из которого вытекает р.Пасвик, распространена в настоящее время по всей системе реки. Ярко выраженный планктонофаг с более эффективным цедильным аппаратом, ряпушка, безусловно, выигрывает в конкуренции со среднетычинковым сигом (Решетников, 1980; Svardson, 1976; Nilsson, 1979). Она активно занимает экологическую нишу среднетычинковых сигов. Резкое увеличение ее численности создает напряженность в пищевой обеспеченности этой формы сига, что является дополнительным стрессовым фактором к уже существующим, обусловленным загрязнением тяжелыми металлами (их влияние будет рассмотрено ниже). Происходит вытеснение среднетычинковых сигов из пелагической зоны в профундальную и литоральную. Переходя на питание бентосными и воздушными организмами, среднетычинковые сиги создают конкуренцию малотычинковым. Таким образом, вторжение ряпушки оказывает косвенное отрицательное влияние и на малотычинковых сигов. Поэтому нет полной ясности в дальнейшей судьбе этих двух форм сига. Скорее всего, малотычинковый сиг, как более приспособленный бентофаг, выдержит конкуренцию со стороны среднетычинковых сигов, но, учитывая короткий цикл развития последних, эта конкуренция может быть очень жесткой. В любом случае, происходит снижение пищевых ресурсов, ранее доступных малотычинковым сигам. Так как эти процессы протекают на фоне значительной техногенной нагрузки, судьба популяций сигов, и в первую очередь среднетычинковых, представляется проблематичной. Ряпушка же прочно обосновалась в пелагиальной зоне, и ее численность будет определяться темпами воспроизводства (естественное и за счет миграции из оз.Инари, зависит от гидрологических условий), колебаниями биомассы зоопланктона и прессом хищников.
Увеличение численности ряпушки и вытеснение среднетычинкового сига в литоральную и профундальные зоны на фоне снижения техногенной нагрузки создали благоприятные условия для роста численности хищных видов. Прежде всего, это кумжа и окунь. Однако следует учитывать загрязнение среды обитания тяжелыми металлами. В этих условиях естественное воспроизводство кумжи проблематично, и ее численность в водоемах системы р.Пасвик будет определяться деятельностью рыбоводных заводов. Увеличение численности окуня наблюдается в оз.Ваггетем (рис.20). Как известно, окунь в процессе онтогенеза занимает три пищевых ниши: младшие возрастные группы питаются зоопланктоном, особи средних размеров потребляют донные организмы, старшие рыбы становятся хищниками (Жаков, 1984). Увеличение численности окуня создает дополнительное напряжение в обеспеченности сигов пищевыми ресурсами. Изменения в структуре рыбной части сообщества изученных водоемов и их прогноз представлены на рис.20.
Другим примером значительных перестроек структуры рыбной части населения могут быть водоемы бассейна р.Поной (центральная часть Кольского п-ова), где отсутствует прямое влияние на водоемы процессов промышленного производства, сельского хозяйства и пр. Здесь в последние годы отмечено
значительное увеличение численности плотвы и язя. Язь, ранее распространенный лишь в верховьях бассейна р.Поной вплоть до оз.Вудьявр (Галкин, 1966; Сурков, 1966), по сообщениям местных жителей, в настоящее время широко обитает как в верхнем, так и в нижнем течении реки. Интенсивному распространению данных видов, по-видимому, способствуют изменения гидрологического и температурного режимов водоемов системы р.Поной, сопровождающиеся массовым развитием высшей водной растительности. На примере малых озер Северо-Запада было показано, что одним из путей сукцессий озер при их зарастании является доминирование плотвы и вытеснение ею остальных видов вплоть до формирования одновидового сообщества (Жаков, 1984).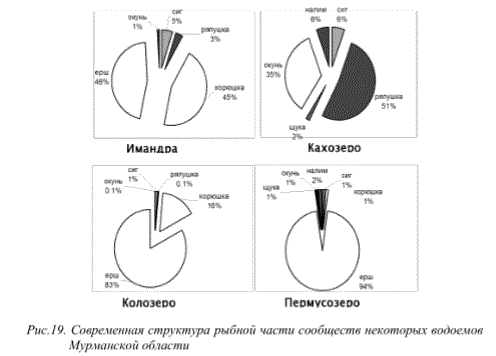
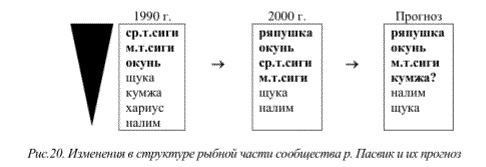
Рис. 20. Изменения в структуре рыбной части сообщества р. Пасвик и их прогноз
Таким образом, несмотря на то, что процесс загрязнения поверхностных вод Мурманской области длится уже не одно десятилетие, мы должны констатировать, что в настоящее время наблюдаются глубокие структурно-функциональные перестройки пресноводных экосистем. Если в прошлом веке интенсивное промышленное загрязнение водоемов отражалось большей частью на состоянии организмов и популяций рыб, то в настоящее время происходит быстрая смена структуры рыбной части сообщества. Значительное снижение качества вод субарктических водоемов, связанное с влиянием многофакторного промышленного загрязнения, интенсификация процессов эвтрофирования водоемов в условиях региональных климатических изменений приводят к развитию не свойственных для Крайнего Севера явлений. В типичных олиготрофных водоемах Субарктики отмечаются серьезные изменения рыбной части сообществ, свидетельствующие о стремительных преобразованиях их трофического статуса на фоне сохраняющегося или повышающегося уровня сублетальной токсичности вод.
Заключение
Интенсивное развитие промышленности на Кольском полуострове в XX веке, разведка и освоение новых запасов полезных ископаемых в последние десятилетия, наряду с климатическими изменениями, привели к возрастающему воздействию на ключевые биологические, геохимические и физические процессы в пресноводных экосистемах Севера. Совместное действие этих факторов вызывает глубокие перестройки в арктических пресноводных экосистемах, снижающие их устойчивость и, в конечном итоге, социально-экономическую значимость. Изменяются скорости и направления сукцессий, интенсивность продукционных процессов, видовой состав и структура сообществ гидробионтов (рис.21).
Начало деградации различных Уровень Средняя температура воздуха
уровней организации биосистем техногенной на грузки (в-летнее сглаживание}
Рис. 21. Динамика некоторых показателей окружающей среды и начало радикальных
изменений биологических систем оз.Имандра
Скорость таких изменений в последние годы чрезвычайно возрастает. Явления, которые в ХХ веке казались невероятными, например, регулярное массовое цветение синезеленых водорослей в арктических озерах, в настоящее время происходят регулярно.
Можно выделить основные направления таких изменений:
• увеличение токсичности водной среды вследствие накопления в озерах загрязняющих веществ;
• изменение трофического статуса озер. Усиление темпов эвтрофикации;
• изменение направленности и скорости сукцессий;
• снижение стабильности экосистемы, повышение рисков катастрофических деградационных изменений.
Совместное действие климатических изменений и загрязнения окружающей среды мало изучено и носит сложный характер. Изменения климата, вне зависимости от направленности таких изменений, могут обусловить существенные изменения в важнейших для региона отраслях экономики - энергетике, коммерческом рыболовстве (включая аквакультуру), туризме, - привести к социальной напряженности вследствие ухудшения условий жизни.
Изменения условий окружающей среды, обусловленные изменениями климата и влиянием загрязнения, могут оказать существенное влияние на природную структуру популяций рыб и их пищевых объектов, что проявляется в исчезновении лососевых и сиговых видов рыб, замене их малоценными (корюшковые, окуневые), изменении технологий аквакультуры и т.д. Цветение синезеленых водорослей вызывает массовую гибель рыб и снижает качество питьевой воды. Это может нанести существенный урон данным секторам экономики, ухудшить условия развития региона. В целом происходит снижение ресурсного потенциала поверхностных вод региона. В то же время повышение трофического статуса озер вследствие процессов эвтрофикации может увеличить их рыбопродуктивность, однако для этого необходимо грамотное, научно обоснованное управление водными ресурсами.
Основные последствия изменений климата и загрязнения окружающей среды для водных систем представлены в схеме (рис.22).
Рис. 22. Основные последствия изменений климата и загрязнения окружающей среды для водных систем
Важным аспектом изучения последствий загрязнений водоемов является оценка потенциальных рисков загрязнения окружающей среды для здоровья населения. Функционирование водных экосистем в этих новых условиях заставляет пересматривать методологические подходы к оценке качества вод и состояния водных ресурсов, а также к организации системы гидроэкологического мониторинга в регионе.
Реализуемый Институтом проблем промышленной экологии Севера КНЦ РАН комплексный подход к исследованию водоемов позволяет получить объективное представление о процессах трансформации пресноводных экосистем под воздействием природных и антропогенных факторов, что, в свою очередь, является научной основой нормирования техногенных нагрузок и прогноза развития природных комплексов.
Н.А.Кашулин, Д.Б.Денисов, С.А.Валькова, О.И.Вандыш, П.М.Терентьев
Список литературы
Алимов А.Ф. Введение в продукционную гидробиологию. Л.: Гидрометиздат, 1989. 151 с.
Алимов А.Ф. Динамика биомассы, продуктивность экосистем континентальных водоемов // Журн. общ. биол. 1997. Т.58, № 3. С. 27-42.
Андроникова И.Н. Классификация озер по уровню биологической продуктивности // Теоретические вопросы классификации озер. СПб.: Наука, 1993.С. 51-72.
Андроникова И.Н. Структурно-функциональная организация зоопланктона озерных экосистем разных трофических типов. СПб.: Наука, 1996. 189 с.
Андроникова И.Н. Зоны экологического риска в прибрежных районах Ладожского озера // Биология внутренних вод. 2007. № 2. С. 3-10.
Антропогенное эвтрофирование озер. М.: Наука, 1976. 200 с.
Аршаница Н.М. Патологоморфологический анализ состояния рыб в полевых и экспериментальных токсикологических исследованиях / Н.М.Аршаница, Л.А.Лесников // Методы ихтиотоксикологических исследований. Л.: ГосНИОРХ; НПО Промрыбвод, 1987. С. 7-9.
Аршаница Н.М. Ихтиологическое состояние водоемов Северо-Запада СССР // Рыб. хоз-во. 1991. № 6. С. 52-54.
Атлас пресноводных рыб России. В 2 т. М.: Наука, 2003. Т. 1. 379 с.; Т. 2. 253 с.
Балушкина Е.В. Функциональное значение личинок хирономид в континентальных водоемах. Л.: Наука, 1987. 179 с.
Баранов И.В. Лимнологические типы озер СССР. Л., 1962. 276 с.
Баринова С.С. Атлас водорослей-индикаторов сапробности (российский Дальний Восток) / С.С.Баринова, ЛА.Медведева. Владивосток: Дальнаука, 1996. 364 с.
Баринова С.С. Биоразнообразие водорослей-индикаторов окружающей среды / С.С.Баринова и др. Тель Авив: PiliesStudio, 2006. 498 с.
Берг Л. С. Рыбы Кольского полуострова. / Л.С.Берг, И.Ф.Правдин // Изв. ВНИОРХ. Л., 1948. Т.ХХУ1, вып.2. 267 с.
Бигон М. Экология. Особи, популяции и сообщества: пер. с англ. В 2 т. / М.Бигон и др. М.: Мир, 1989. Т.2. 477 с.
Биологическая продуктивность северных озер. Озера Зеленецкое и Акулькино / под ред. Винберга. Л., 1975. Т.57. 182 с.
Болотова Н.Л. Изменения рыбной части сообщества и уловов при эвтрофировании крупного северного озера / Н.Л.Болотова и др. // Вопросы ихтиологии. 1996. Т.36, № 4. С. 470-480.
Большие озера Кольского полуострова / под ред. Л.Ф. Форш, В.Г. Драбковой. Л.: Наука, 1975. 350 с.
Валькова С.А. Особенности бентосных сообществ губы Монче (оз.Имандра, Кольский полуостров) // Современные проблемы гидроэкологии: тез. докл. 4-й Междунар. науч. конф. (Санкт-Петербург, 11-15 октября 2010 г.) / ЗИН РАН. СПб., 2010. С. 33.
Васнецов В.В. О закономерностях роста рыб // Очерки по общим вопросам ихтиологии. М.-Л.: Изд-во Акад. наук СССР, 1953. С. 218-226.
Васнецов В.В. Этапы развития костистых рыб // Очерки по общим вопросам ихтиологии. М.-Л.: Изд-во Акад. наук СССР, 1953. С. 207-217.
Волхонская Н.И. Зоопланктон Ковдозерского водохранилища // Рыбы Мурманской области. Мурманск, 1966. 336 с.
Галкин Г.Г. Ихтиофауна водохранилищ и озер Мурманской области / Г.Г.Галкин и др. // Рыбы Мурманской области. Условия обитания, жизнь и промысел. Мурманск: Изд-во ПИНРО, 1966. С. 177-193.
Давыдова Н.Н. Диатомеи в донных отложениях (Сейдозеро) // Большие озера Кольского полуострова. Л., 1971. С.140-168.
Денисов Д.Б. Изменения гидрохимического состава и диатомовой флоры донных отложений в зоне воздействия горнорудного производства (Кольский полуостров) // Водные ресурсы. 2007. Т.34, № 6. С. 719-730.
Денисов Д.Б. Экологические особенности функционирования разнотипных субарктических водоемов / Д.Б.Денисов, Н.А.Кашулин. 2007. ИКЬ: http://www.kolasc.net.ru/russian/sever07/sever07_1.pdf.
Денисов Д.Б. Динамика водорослевых сообществ горных субарктических водоемов // Экологические проблемы северных регионов и пути их решения: материалы всерос. науч. конф. с междунар. участием (14-16 октября 2008 г.). Апатиты: Изд. КНЦ РАН, 2008. Ч.І. С. 205-210.
Денисов Д.Б. Водорослевые сообщества различных ландшафтов Кольского Севера в оценке состояния водных экосистем // Водоросли: проблемы таксономии, экологии и использование в мониторинге: материалы II Всерос. конф. (Сыктывкар, 5-9 октября) / Ин-т биологии Коми НЦ УрО РАН. Сыктывкар, 2009. С. 270-272.
Денисов Д.Б. Экологические особенности водорослевых сообществ разнотипных субарктических водоемов // Вестник Кольского научного центра РАН. 2010. № 1. С. 48-55.
Деньгина Р.С. Зоопланктон и зообентос озера Имандра // Экосистема озера Имандра под влиянием техногенного загрязнения. Апатиты: Изд. Кольского филиала АН СССР, 1980. С. 78-115.
Дольник Т.В. Зоопланктон и зообентос / Т.В.Дольник, Г.А.Стальмакова // Большие озера Кольского полуострова. Л., 1975. С. 317-336.
Дубровина Л.В. К вопросу о влиянии биотических и абиотических факторов среды на токсичность тяжелых металлов / Л.В.Дубровина и др. // Тез. докл. II Всесоюз. конф. по рыбохоз. токсикологии. СПб., 1991. Т.1. С. 168-170.
Жадин В.И. Реки, озера и водохранилища СССР, их фауна и флора /
В.И.Жадин, С.В.Герд. М., 1961. 600 с.
Жаков Л.А. Формирование и структура рыбного населения озер Северо-Запада СССР. М., 1984. 144 с.
Жизнеспособность популяций: Природоохранные аспекты. М.: Мир, 1989. 224 с.
Зверева О С. Бентос и общие вопросы гидробиологии Вашуткиных озер // Гидробиологическое изучение и рыбохозяйственное освоение озер Крайнего Севера СССР. М., 1966. С. 112-137.
Здановски Б. Оценка изменений, вызванных сбросом теплых вод в систему Конинских озер // Биологические ресурсы водоемов Балтийского моря: тез. докл. XXIII науч. конф. по изучению водоемов Прибалтики. Петрозаводск, 1991. С. 205-206.
Зинова А.Д. Сравнительная характеристика исследованных озерно-речных систем Монче- и Волчьей тундр / А.Д.Зинова, А.А.Нагель // Тр. отдел. гидрол. ленингр. обл. гидрометеорол. упр. Л., 1935. Т.І. С. 113-132.
Ильяшук Б.П. Реликтовые ракообразные в условиях длительного загрязнения субарктического оз.Имандра (результаты наблюдений за период 1930-1998 гг.) // Экология. 2002. № 3. С. 215-219.
Каган Л.Я. Исследования диатомей на Кольском полуострове в XX веке / Л.Я.Каган, Д.Б.Денисов // Морфология, экология и биогеография диатомовых водорослей: сб. тез. VIII Школы диатомологов России и стран СНГ / под ред.
С.И.Генкала; Ин-т биологии внутренних вод им. И.Д.Папанина. Борок, 2002. С. 38-39.
Калюжин С.М. Атлантический лосось Белого моря: проблемы
воспроизводства и эксплуатации. Петрозаводск: ПетроПресс, 2003. 264 с.
Канаев А.И. Ветеринарная санитария в рыбоводстве. М.: Агропромиздат, 1985. 280 с.
Канеп С.В. Взаимосвязь энергетического обмена, потенциальных размеров, скорости роста и полового созревания у сиговых рыб: Второе всесоюз. совещ. по биологии и биотехнике разведения сиговых рыб: тез. докл. Петрозаводск, 1981. С. 9-11.
Карамушко О.В. Ихтиофауна пресных вод Мурмана / О.В.Карамушко, Е.Г.Берестовский // Ихтиофауна малых рек и озер Восточного Мурмана: биология, экология, биоресурсы. Апатиты: Изд. Кольского науч. центра РАН, 2005. С. 36-42.
Кашулин Н.А. Рыбы пресных вод Субарктики как биоиндикаторы техногенного загрязнения / Н.А.Кашулин, А.А.Лукин, П.-А.Амундсен. Апатиты: Изд. Кольского науч. центра РАН, 1999. 142 с.
Кашулин Н.А. Рыбы малых озер Северной Фенноскандии в условиях аэротехногенного загрязнения. Апатиты: Изд. Кольского науч. центра РАН, 2004. 130 с.
Кашулин Н.А. Современные подходы к оценке процессов трансформации пресноводных экосистем Севера / Н.А.Кашулин и др. // Сб. науч. трудов Кольского науч. центра РАН. Апатиты: Изд. КНЦ РАН, 2005. Т.1. С. 337-346.
Кашулин Н.А. Антропогенные изменения водных систем Хибинского горного массива (Мурманская область) / Н.А.Кашулин и др. Апатиты: Изд. Кольского науч. центра РАН, 2008. Т.1. 250 с.
Кашулин Н.А. Экологический каталог озер Мурманской области. Северо-западная часть Мурманской области и приграничные территории сопредельных стран. В 2 ч. / Н.А.Кашулин и др. Апатиты: Изд. Кольского науч. центра РАН, 2009. Ч.1. 226 с.; Ч.2. 262 с.
Китаев С.П. Экологические основы биопродуктивности озер разных природных зон. М., 1984. 207 с.
Комулайнен С.Ф. Методические рекомендации по изучению фитоперифитона в малых реках. Петрозаводск: Карельский науч. центр РАН, 2003. 43 с.
Комулайнен С.Ф. Библиография работ по водорослям Европейского Севера России (Республика Карелия, Мурманская область) / С.Ф.Комулайнен и др. Петрозаводск: Карельский науч. центр РАН, 2006. 66 с.
Комулайнен С.Ф. Альгологические исследования в озерно-речных системах Севера европейской части России // Альгология. 2007. Т.17, № 2. С. 220-229.
Королева И.М. Влияние загрязнения на морфофизиологические показатели сигов Coregonus Їауагеїш в водоемах Кольского Севера: дис. ... канд. биол. наук. Апатиты, 2001. 186 с.
Кошелев Б.В. Изучение закономерностей репродуктивного процесса у рыб // Экологические аспекты исследований водоемов-охладителей АЭС. М.: ИЭМЭЖ АН СССР, 1983. С. 54-66.
Крючков В.В. Экология водоемов-охладителей в условиях Заполярья / В.В.Крючков, Т.И.Моисеенко, В.А.Яковлев. Апатиты: Изд. Кольского филиала АН СССР, 1985. 132 с.
Кудерский Л.А. Динамика стад промысловых рыб внутренних водоемов. М.: Наука, 1991. 151 с.
Лапин В.И. Закономерности внутривидовой изменчивости обменных процессов и характера воспроизводства у рыб / В.И.Лапин, Н.Н.Лапина, М.И.Шатуновский // Особенности репродуктивных циклов у рыб в водоемах разных широт. М.: Наука, 1985. С. 65-77.
Летанская Г.И. Фитопланктон и первичная продукция озер Кольского полуострова // Озера различных ландшафтов Кольского полуострова. Л.: Наука, 1974. Ч.2. С. 78-119.
Лукин А.А. Патологии рыб как индикатор качества вод Кольского Севера // Проблемы химического и биологического мониторинга экологического состояния водных объектов Кольского Севера. Апатиты: Изд. КНЦ РАН, 1995. С. 105-119.
Лукин А.А. Интродукция радужной форели Рагазо1шо ту1а88 в озеро Имандра (Кольский полуостров) // Вопросы ихтиологии. 1998. Т.3, № 4. С. 485-491.
Лукьяненко В.И. Экологические аспекты ихтиотоксикологии. М.: Агропромиздат, 1987. 240 с.
Макарцева Е.С. Зоопланктон озер различных ландшафтов Кольского полуострова // Озера различных ландшафтов Кольского полуострова. Л., 1974. Ч.2. С. 143-179.
Макрушин А.В. Биоиндикация загрязнения внутренних водоемов // Биологические методы оценки природных вод. М.: Наука, 1984. С. 123-137.
Методы биоиндикации и биотестирования природных вод. Л.: Гидрометеоиздат, 1989. 277 с.
Моисеенко Т.И. Изменение физиологических показателей рыб как индикатор качества водной среды // Мониторинг природной среды Кольского Севера. Апатиты: Изд. КНЦ РАН, 1984. С. 51-57.
Моисеенко Т.И. Антропогенные преобразования водных экосистем Кольского Севера / Т.И.Моисеенко, В.А.Яковлев. Л.: Наука, 1990. 221 с.
Моисеенко Т.И. Сиг как тест-объект для биоиндикации качества вод озер Крайнего Севера / Т.И.Моисеенко, А.А.Лукин, Н.А.Кашулин // Современные проблемы сиговых рыб. Владивосток, 1991. С. 213-224.
Моисеенко Т.И. Теоретические основы нормирования антропогенных нагрузок на водоемы Субарктики. Апатиты: Изд. КНЦ РАН, 1997. 261 с.
Моисеенко Т.И. Изменение стратегии жизненного цикла рыб под воздействием хронического загрязнения вод // Экология. 2002. № 1. С. 50-60.
Моисеенко Т.И. Водная экотоксикология: Теоретические и прикладные аспекты / Т.И.Моисеенко; Ин-т водных проблем РАН. М.: Наука, 2009. 400 с.
Моисеенко Т.И. Антропогенная трансформация арктической экосистемы озера Имандра: тенденции к восстановлению после длительного периода загрязнения / Т.И.Моисеенко и др. // Водные ресурсы. 2009. Т.36, № 3. С. 312-325.
Мордухай-Болтовской Ф.Д. Проблема влияния тепловых и атомных электростанций на гидробиологический режим водоемов // Экология организмов водохранилищ-охладителей. Л.: Наука, 1975. С. 7-69.
Муравейко В.М. Стальноголовый лосось в реках Восточного Мурмана / В.М.Муравейко и др. // Виды-вселенцы в европейских морях России. Апатиты: Изд. КНЦ РАН, 2000. С. 269-272.
Нилова О.И. Гидробиологическая характеристика р. Поноя и ее притоков // Рыбы Мурманской области. Мурманск: Кн. изд-во,1966. С. 105-111.
Озера различных ландшафтов Кольского полуострова. Л., 1974. Ч.2. С. 319.
Петровская М.В. Характеристика зоопланктона озер Мурманской области // Рыбы Мурманской области. Мурманск, 1966. С. 84-90.
Порецкий В.С. Диатомовые Кольского полуострова в связи с микроскопическим составом кольских диатомитов / В.С.Порецкий, А.П.Жузе, В.С.Шешукова // Труды Геоморф. ин-та АН СССР. 1934. Т.8. С. 95-200.
Решетников Ю.С. Экология и систематика сиговых рыб. М.: Наука, 1980. 300 с.
Решетников Ю.С. Изменение структуры рыбного населения эвтрофируемого водоема / Ю.С.Решетников, О.А.Попова, О.П.Стерлигова. М.: Наука, 1982. 234 с.
Решетников Ю.С. Время наступления половой зрелости / Ю.С.Решетников и др. // Пелядь Coregonus peled (Ошєііп, 1788). Систематика, морфология, экология, продуктивность. М.: Наука, 1989. С. 161-167.
Решетников Ю.С. Влияние нового вида-вселенца на экосистему (на примере ряпушки реки Пасвик) / Ю.С.Решетников, О.А.Попова, П.-А.Амундсен // Водные экосистемы: трофические уровни и проблемы поддержания биоразнообразия: материалы Всерос. конф. с междунар. участием. Вологда, 2008. С. 347-350.
Руководство по методам гидробиологического анализа поверхностных вод и донных отложений / отв. ред. В.А.Абакумов и др. СПб.: Гидрометеоиздат, 1992. 305 с.
Рыбы Мурманской области. Мурманск, 1966. 334 с.
Рылов В.М. Материалы к фауне свободноживущих пресноводных Copepoda северной России. Ч.1: Calanoida и Cyclopoida // Ежегодник Зоол. музея Российской академии наук. 1922. Т.22. С. 247-310.
Сажин Е.В. Популяция плотвы (Rutilus тМш) Кубенского озера // Водные экосистемы: трофические уровни и проблемы поддержания биоразнообразия: материалы Всерос. конф. с междунар. участием. Вологда, 2008. С. 364-366.
Семенова А.С. Изменение показателей зоопланктона Куршского залива в период «гиперцветения» синезеленых водорослей // Вода: химия и экология. 2009. № 9. С. 2-6.
Семенович Н.И. Гидрологические исследования озера Имандра в 1930 г. // Материалы к изучению водоемов Кольского полуострова. Мурманск, 1940. Сб.1. С. 151-191.
Сурков С.С. Общая характеристика особенностей видового состава ихтиофауны Мурманской области // Рыбы Мурманской области. Условия обитания, жизнь и промысел. Мурманск: Изд-во ПИНРО, 1966. С. 147-151.
Телеш И.В. Роль планктонных ракообразных в водных экосистемах разного типа (на примере Ладожского озера, р.Невы и Невской губы) // Материалы VII съезда гидробиол. о-ва РАН. Казань, 1996. Т.2. С. 90-92.
Терентьев П.М. Особенности динамики популяций рыб в водоемах Кольского Севера в условиях их аэротехногенного загрязнения: автореф. дис. ... канд. биол. наук. Петрозаводск, 2005. 28 с.
Чинарева И.Д. Патогистологические изменения, встречающиеся у рыб бассейна Ладожского озера // Сб. науч. трудов ГосНИОРХ. 1988. С. 24-32.
Шаров А.Н. Фитопланктон водоемов Кольского полуострова. Петрозаводск: Карельский науч. центр РАН, 2004. 113 с.
Шарова Ю.Н. Особенности функционирования системы воспроизводства рыб Кольского Севера в условиях техногенного загрязнения: автореф. дис. ... канд. биол. наук. Петрозаводск, 2000. 26 с.
Шилов И.А. Физиологическая экология животных. М.: Высш. шк., 1985. 328 с.
Шитиков В.К. Количественная гидроэкология: методы современной идентификации / В.К.Шитиков, Г.С.Розенберг, Т.Д.Зинченко. Тольятти: ИЭВБ РАН, 2003. 463 с.



