Продукционный цикл фитопланктона
Основными факторами, определяющими первичную продукцию фитопланктона, являются толщина фотического слоя, обеспеченность фитопланктона биогенными элементами и продолжительность продукционного цикла.
В целом указанные параметры зависят от конкретных климатических и гидрологических условий каждого района и проявляются в сроках наступления весеннего развития, которые тесно связаны следовым режимом и окончания продукционного цикла; кроме того, эти параметры определяются запасом биогенных элементов, образовавшимся в фотическом слоек началу весеннего развития и определяемым речным стоком и глубиной зимнего конвективного перемешивания, летней вертикальной турбулентной диффузией, количеством солнечной энергии (зависящей в свою очередь от времени года и климатических факторов), а также структурно-функциональными характеристиками фитопланктона.
Фитопланктон Белого моря представляет собой качественно обедненный планктон Баренцева моря с большой примесью «литоральных» эпифитных и пресноводных форм (Кокин, Кольцова, 1972). В Белом море по сравнению с южной частью Баренцева моря по числу видов преобладают холодолюбивые диатомеи и снижается видовое разнообразие перидиней. Подоле содержания водорослей в составе фитопланктона (63%) Белое море приближается к таким высокоарктическим эпиконтинентальным морям, как Карское (67%) и Лаптевых (64 %), что лишний раз указывает на его арктический характер. Формирование качественного и количественного состава фитопланктона в различные биологические сезоны происходит под влиянием как биотических, так и абиотических факторов среды.
Изменение структурных и функциональных показателей сообщества в течение продукционного цикла можно проследить на примере анализа биологических сезонов в Кандалакшском заливе в 1970 г. (Федоров, Бобров, 1977, Бобров, 1982).
Всего за период наблюдений с 15 апреля по 26 октября в 1970 г. поставлена 51 станция (дважды в неделю). Продукцию определили радиоуглеродным методом склянок на горизонтах 0,2.5, 5, 10, 25, 50, 75 м и у дна. Суммарную солнечную радиацию измеряли пиранометром Янышевского в подводной модификации. Одновременно с постановкой опытов по определению первичной продукции с соответствующих горизонтов проводили батометрические количественные сборы фитопланктона и изучение некоторых гидролого-гидрохимических параметров. При расчете эффективности фотосинтетического процесса данные измерения первичной продукции были выражены в энергетических единицах, исходя из следующего расчета: на фиксацию I моля CO2 расходуется 112 калорий световой энергии, а эта величина соответствует 9.36 кал/мгC.
Переход от численности особей фитопланктона к весовым характеристикам производили по формуле:
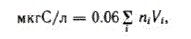
Формула получена с учетом того, что в сухой биомассе водорослей содержится 35 % золы (Федоров и др., 1974).
Величины суточных сумм радиации в течение вегетационного периода 1970 г. колебались от 20.6 до 2300 кал/(см". день). Увеличение проникающей в BOAy солнечной радиации в первой декаде мая до 100.6—181.4 кал/(см" - день) совпадает повремени с вскрытиемледового покрова. Некоторое снижение значений суммарной солнечной энергии было вызванометеорологическими условиями Улучшение погодных условий и увеличение длины светового дня привело к практическому совпадению светового максимума (2300 кал/(см". день)) с календарным днем летнего солнцестояния. Последующее падение дневных сумм солнечной радиации имеет довольно плавный характер и обусловливается уменьLEHI HEM продолжительности CBPTOBOTO IH H H BECOTE CONHL kl. К концу вегетационного сезона величина солнечной энергии падает до 20 кал/(см". день).
Таким образом, за весь период наблюдений, охватывающий 183 дня, величина энергии выразилась суммарным значением 22.8 ккал/см". Неучтенная энергия проникающей солнечной радиации в период осении «подледной весны» была получена экстраполяцией и составила 22 ккал/см". Всего за сезон в водоем проникло 25 ккал/см". Последнее значение можно считать годовой суммой, поскольку в зимние месяцы количество энергии солнечной радиации, проникающей под ледяной покров, незначительно, Изучение распространения солнечной энергии в толще воды на протяжении вегетационного периода позволило рассчитать коэффициент поглощения света водой k=0.37-002.
Температура в поверхностном горизонте в течение вегетационного сезона колебалась от — 1.54 до 15.65 °С. С глубиной температурные колебания сглаживались. Весенний период после вскрытия льда характеризуется нарушением гомотермии и установлением отчетливо выраженной стратификации. Зона температурного скачка с ростом проникающей солнечной энергии и соответствующим прогревом ВМ постепенно перемещалась из поверхностного в более глубокие слои. В период максимального значения солнечной энергии термоклин располагался на глубине 25 м и удерживался на этом горизонте до конца сентября. В октябре температурный скачок был расположен глубже и обнаруживалась тенденция к гометермии.
В табл. 5.1 представлены некоторые абиотические характеристики Белого моря, полученные на основе анализа их сезонной LHHaMHK.H.
Анализ вегетационного развития планктона и его продукционHEIM Xapa KTepH cTH K позволяет выделить и установить продолжительность основных периодов вегетации, названных В. Г. Богоровым (1938) биологическими весной, летом и осенью. Для Белого моря характерны все три периода. Их продолжительность для Кандалакшского залива в 1970 г. составила 48, 92 и 43 сут соответственно.

Результаты, характеризующие абсолютные величины продукционного процесса в различные периоды вегетации, представлены в табл. 5.2. Коэффициент Р/Сент рассчитывали как отношение первичной продукции к биомассе (выраженной в углероде) для Bice TO CTO,JI (5a B OIE bil.
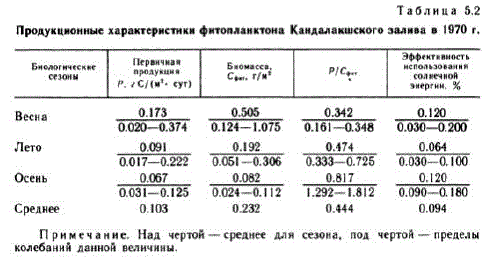
Примечание. Над чертой — среднее для сезона, под чертой — пределы колебаний данной величины.
Начало биологической весны отмечено впервой декаде апреля, Весенний подледный климакс (Федоров, 1970a; Кокин, Кольцова, 1972) продолжается около 2 недель и характеризуется устойчивым P/Cфи-коэффициентом. Распределение экониш водорослей соответствует второй модели Мак-Артура (Кокин, Кольцова, 1972).
Весенняя вспышка величин биомассы и численности водорослей начинается подо льдом и после вскрытия льда достигает максимума. Фитопланктон представлен BH La MH JULHETOMOBOTO KOM TIJEKCE: Fragilaria oceanica, Chaetoceros holsa ficus, Nitzschia frigida, Maticula septentrionaliа. Самым массовым видом была F. oceапіса. Фотический слой обогащен биогенными элементами и содержит 86 % всей биомассы фитопланктона, которая, так же как и скорость фотосинтеза, в этот период достигает максимального значения за весь вегетационный период (см. табл. 5.2). Выедание водорослей зоопланктоном (Конопля, 1973) приводит к падению биомассы водорослей и снижению пищевой конкуренции за биогенные элементы, при этом распределение водорослей соответствует уже первой модели Мак-Артура.
Летний максимум развития фитопланктона (первый), обусловленный увеличением числа диатомовых, после спада их численности «подхватывается» перидиниевыми водорослями, образующими второй летний максимум. С середины июля и до конца августа устанавливается летний климакс с устойчивым P/Cфиткоэффициентом. Доминирующими формами биологического лета оказываются Sceletoпeтa costalит и виды рода Chaetoceros (главным образом Ch. сотргеssиs). Первичная продукция в этот период характеризуется средней величиной 0,091 г С/(м", сут), а биомасса — 0.192 гC/м". Развита трофическая конкуренция за биогенные элементы,
Осенняя вспышка фитопланктона выражена слабо. Она наблюдается сконца августа до середины сентября и поросту биомассы ee, Rb Ae/y!"b, "pyAhQ. Только увеличение первичной продукции до 0.125 г С/(м", сут) и Р/Сеит-коэффициента (до 1812) в 20-х числах сентября указывает на кратковременную активацию фитопланктона. В этот период наблюдается развитие Dinophyes пorwegica — холодолюбивого доминанта биологической осени (Федоров, 1969). Период осенней сукцессии приводит экосистему к кратковременному климаксу, который длится вплоть до конца сентября. Дальнейшее снижение температуры и солнечной радиации, а также увеличение слоя перемешивания в связи с нарушением скачка плотности приводит к перераспределению биомассы фитопланктона по глубине и преобладанию процессов деструкции над продукцией.
В целом такой ход продукционного цикла фитопланктона можно ожидать и в других районах моря. Возможные различия будут выражены в сроках начала и конца вегетации фитопланктона, а также в некоторых различиях качественного состава доминирующих видов водорослей (Житина, 1981), что несомненно отразится на результатах продукционных процессов фитопланктона этих частей акватории.
Данные исследований продукционного цикла фитопланктона в 1970 г. позволили уточнить классификацию трофности Белого моря. В качестве дополнительной характеристики была использована эффективность использования солнечной энергии фитопланктоном (см. табл. 5.2), являющаяся интегральной характеристикой состояния фитопланктона, зависящая от комплекса условий окружающей среды и не обнаруживающая прямой связи с биомассой фитопланктона (Федоров, Бобров, 1977).
Если сравнить расчетные значения основных продукционных характеристик фитопланктона с известными данными для различных по продуктивности районов океана (Сорокин, 1973), то можно сделать вывод, что Кандалакшский залив относится к малопродуктивным районам с уровнем продукции фитопланктона 0,05— 0.10 г С/(м*- cут) и эффективностью использования солнечной энергии 0.04 %, характерной для олиготрофных вод. Следовательно, Кандалакшский залив, согласно классификации О. И. Кобленц-Мишке (Кобленц-Мишке и др., 1970), можно отнести к 1-му классу трофности с очень незначительным перемешиванием Глубинных вод.



