3оопланктон
Исследования зоопланктона Белого моря, начавшиеся в конце XIX столетия, первое время носили лишь качественный характер (Вагнер, 1885; Педашенко, 1897; Линко, 1900). Они продолжались и в дальнейшем (Виркетис, 1928; Хмызникова, 1947 и др.), но, начиная с 30–40-х годов прошлого столетия (Богоров, 1932; Яшнов, 1940; Хмызникова, 1947 и др.), к ним стали добавляться количественные данные о показателях обилия, распределении и жизненных циклах зоопланктонных организмов (Камшилов, 1951, 1952, 1957).
Эта составляющая планктонологических работ стала особенно ощутимой после начала исследований на биологических станциях Московского университета и Зоологического института РАН (Перцова, 1962, 1970; Прыгункова, 1967, 1974, 1987а; Перцова, Прыгункова, 1995).
В результате этих работ было установлено, что видовой состав беломорского зоопланктона значительно беднее такового в Баренцевом море. Основу планктонной фауны составляют Сopepoda, которых по разным источникам насчитывается около 35 видов. Образуя большую часть биомассы зоопланктона, эти ракообразные служат главным источником пищи для многих обитателей пелагиали Белого моря (рыб, медуз и гребневиков).
Наиболее разнообразен по видовому составу зоопланктон Кандалакшского залива. Минимальное число видов зоопланктонных организмов было зарегистрировано в Мезенском заливе и Воронке.
С зоогеографической точки зрения основу планктона составляют арктическо-бореальные и бореальные элементы. Последние в массе развиваются в летнее время в поверхностном, хорошо прогреваемом горизонте. При этом скорость их развития близка или даже превосходит таковую в бореальных районах Мирового океана, где эти рачки дают за год до 6 поколений (Marshall, 1949). Некоторые виды данного комплекса, например, Oithona similis, в отдельные годы могут иметь в Белом море 3 поколения (Прыгункова, 1974). Арктические виды большую часть года проводят на глубинах более 25 м, поднимаясь к поверхности лишь весной и в начале лета, когда происходит их активное развитие. Затем они вновь опускаются на глубину, а их развитие останавливается. Арктическо-бореальные формы, отличающиеся наибольшей эвритермностью, развиваются в течение длительного периода.
Динамика развития зоопланктона меняется от года к году в зависимости от температуры воды. Обычно изменения обилия носят моноциклический характер и приходятся на июль – август. В наиболее теплые годы наблюдаются два пика численности: первый приходится на начало июня, а второй – на август (рис. 16).
В холодные годы обилие зоопланктона нарастает крайне медленно и достигает своего единственного максимума лишь в конце августа. При этом пик обилия менее выражен, чем в более теплые годы. Чередование теплых и холодных лет и соответствующих им изменений обилия зоопланктона происходит с интервалом в 5–6 лет (Бабков, Прыгункова, 1974; Прыгункова, 1987; Перцова, 1970, 1980, 1984; Перцова, Прыгункова, 1995).
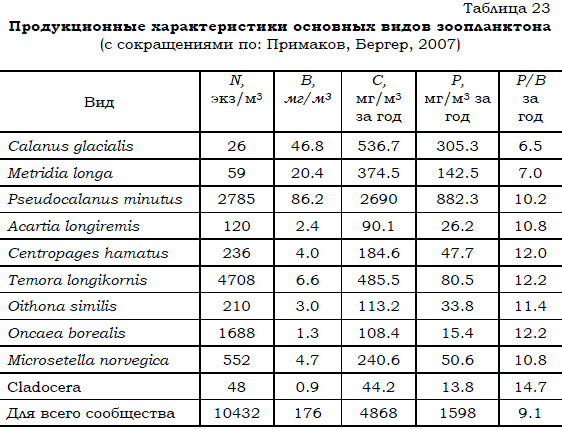
Несмотря на наличие значительных межгодовых колебаний, среднемноголетние показатели обилия сохраняются практически без изменений на одном уровне на протяжении, по крайней мере, 40 лет (рис. 17).
Рис. 17. Многолетние изменения средних значений плотности зоопланктона за летние месяцы (с июля по сентябрь) в устьевой части губы Чупа (по: Бергер и др., 2004).
Жирной линией обозначен полиномиальный тренд третьей степени.
Пространственные изменения обилия беломорского зоопланктона весьма значительны. Если говорить о вертикальных изменениях, то с глубиной биомасса зоопланктона
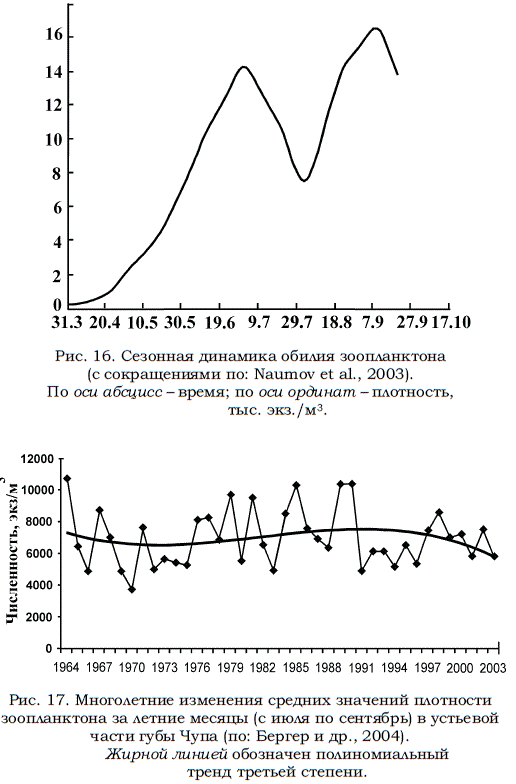
Рис. 16. Сезонная динамика обилия зоопланктона (с сокращениями по: Naumov et al., 2003). По оси абсцисс – время; по оси ординат – плотность, тыс. экз./м3.
значительно уменьшается. Так, например, по данным мониторинга в устьевой части губы Чупа в период с 1963 по 1981 гг. средняя биомасса зоопланктона в летнее время в поверхностных водах (слой 0–10 м) была равна 356 мг/м3 (Berger et al., 2003). В те же годы в центральной части Кандалакшского залива на глубинах от 100 до 300 м она была меньше в 3 с лишним раза: 114 мг/м3 (Прыгункова, 1987).
Наряду с вертикальными градиентами, весьма существенны изменения обилия зоопланктона, регистрируемые в разных районах моря. Как правило, в прибрежных районах, особенно в кутах заливов и губ, зоопланктон значительно обильнее, чем в открытых участках, вдали от берегов. Что касается разных частей акватории моря, то приходится признать, что не все они изучены достаточно подробно. Наиболее исследован зоопланктон Кандалакшского залива. По многолетним данным (с 1963 по 1981 гг.) средняя для всей водной толщи залива биомасса зоопланктона колебалась на глубинах от 0 до 300 м в относительно небольших пределах: от 140 до 232 мг/м3 при среднем значении около 200 мг/м3 (Прыгункова, 1987).
Длительное время (примерно до конца 80-х годов прошлого столетия) количественные сборы зоопланктона в других частях Белого моря (кроме Кандалакшского залива) были единичны (Яшнов, 1940; Эпштейн, 1963; Перцова, 1980; Прыгункова, 1987). Кроме того, полученные данные было трудно сравнивать из-за того, что сборы зоопланктона осуществлялись не по единой программе. Ситуация значительно улучшилась после того, как сотрудники СевПИНРО с 1970 по 1988 гг. осуществили несколько рейсов и исследовали распределение зоопланктона во всей акватории Онежского и Двинского заливов, включая и плохо исследованные до этого кутовые участки этих заливов (Трошков, 1998; Трошков, Слонова, 2000; Трошков, Фролов, 2005). По полученным данным среднегодовая биомасса зоопланктона Онежского залива в изученные годы составляла 150 мг/м3 при колебаниях по сезонам от 190 г/м3 (весна–лето) до 66 мг/м3 (осень). За этот же период наблюдений среднегодовая биомасса зоопланктона Двинского залива составила 295 мг/м3. Весенняя биомасса в среднем была – 278, летняя – 378, а осенняя – 91 мг/м3 (Трошков, Слонова, 2000).
В результате анализа всех имеющихся материалов можно подвести некоторые итоги и охарактеризовать среднее обилие зоопланктона в Белом море. При общей обедненности видового состава беломорского зоопланктона по сравнению с баренцевоморским, о чем уже говорилось в начале этого раздела, количественные показатели его развития не уступают таковым в Баренцевом море. В конце 80-х годов прошлого столетия было определено, что средняя биомасса зоопланктона в Белом море составляет по разным подсчетам от 160 до 200 мг/м3 (Перцова, Прыгункова, 1995). Если учесть результаты исследований последних лет, значительно пополнивших наши представления об обилии зоопланктона Белого моря в целом, то можно подсчитать, что средняя биомасса зоопланктона ближе не к 160, а к 200 мг/м3. Эту величину мы и будем использовать в дальнейших расчетах баланса вещества и энергии в беломорских экосистемах. При этом следует отметить, что как в более богатых районах моря с «бассейновой» структурой вод, так и в более бедных участках акватории, характеризующихся повышенной гидродинамикой и вертикальной гомотермией, были неоднократно зарегистрированы случаи гораздо более высокого обилия зоопланктона. Так, максимальные биомассы зоопланктона, обнаруженные в Кандалакшском заливе, достигали 2.5 г/м3 (Перцова, Прыгункова, 1995), а в Двинском заливе – 0.8 г/м3 (Бондаренко, 1994) и 7.6 г/м3 (Трошков, Слонова, 2000). Наибольшая биомасса зоопланктона, отмеченная в Горле, была равна 1.8 г/м3 (Трошков, 1998).
Что касается зоопланктона Баренцева моря, то согласно современным данным (Дружинина, Марасаева, 2004), его обилие (средняя биомасса) оценивается величинами от 50 до 200 мг/м3. Сравнение этих данных по двум соседним морям в очередной раз убеждает в ошибочности причисления Белого моря к разряду водоемов с низкой биологической продуктивностью. Беломорский зоопланктон не только не беднее баренцевоморского, но даже превосходит последний по показателям обилия.
Таким образом, в давнем споре о том, беднее ли количественно беломорский зоопланктон, чем баренцевоморский (Зенкевич, 1947, 1963; Эпштейн, 1963) или нет (Яшнов, 1940; Камшилов, 1951, 1957), можно, наконец, поставить точку. Очевидно, что полярность взглядов этих исследователей была обусловлена недостаточностью данных об обилии зоопланктона в сравниваемых водоемах. Сейчас, когда этот пробел в основном заполнен, можно утверждать, что зоопланктон Белого моря не беднее количественно (по биомассе) баренцевоморского, хотя и уступает последнему по числу видов.
В связи с этим весьма показателен вывод из уже упоминавшейся работы (Трошков, Слонова, 2000, с.161): «В целом биомассы зоопланктона Двинского и Онежского заливов находятся на достаточно высоком уровне и не только не уступают биомассе баренцевоморского зоопланктона, но в некотором отношении и превосходят ее».
Приведенные выше результаты позволяют сравнить обилие зоопланктона в Белом море и с аналогичными показателями в Мировом океане в целом. Ранее (Berger, Kosobokova, 2001) это уже было сделано. Сейчас, однако, целесообразно повторить сравнение, исходя из уточненных данных. Объем Белого моря (5.4 тыс. км3) примерно в 250 тысяч раз меньше объема Мирового океана (1341 млн. км3 – Моисеев, 1989; Гершанович и др., 1990). Содержание зоопланктона в Мировом океане оценивается величиной порядка 21.109 т (Виноградов, 1955; Богоров и др., 1968). В Белом море при среднем обилии 200 мг/м3, общая биомасса зоопланктона составляет 1.08 .106 т. Следовательно, биомасса зоопланктона Белого моря меньше таковой в Мировом океане приблизительно в 19 тысяч раз.
Из этого следует, что общая масса зоопланктона в Белом море, отнесенная к единице объема воды, превосходит таковую в Мировом океане в 13 раз.
Зоопланктон занимает промежуточное положение в трофической цепи между фитопланктоном, с одной стороны, и рыбами – с другой. Вместе с величиной первичной продукции показатели обилия и продуктивности сообществ зоопланктона являются наиболее адекватными показателями общего продукционного потенциала того или иного водоема (Богоров, 1974).
Задача расчета продукции зоопланктона Белого моря решалась двумя различными способами: во-первых, с помощью балансового равенства обменных процессов в популяциях основных видов планктонных ракообразных (Примаков, Бергер, 2007), и, во-вторых, с помощью P/Bкоэффициента и показателей обилия зоопланктона (Бергер, 2005; Бергер и др., 1995).
В соответствии с классическими представлениями (Boysen-Jensen, 1919; Thienemann, 1931) продукция популяции в соответствии с тем, что уже отмечалось ранее (см. гл. 1), рассматривается (Винберг, 1968) как сумма прироста биомассы с какого-то определенного момента (Bt – B0) и биомассы элиминированных за это время особей (Be):
P = (Bt – B0) + Be.
Это равенство формально обеспечивает расчет величины продукции лишь в самом общем виде. Такой подход применим только к популяциям с четко выраженными сменяющими друг друга генерациями, появляющимися в сжатые по сравнению с длительностью своего существования сроки.
К планктонным популяциям он приложим с большими ограничениями, хотя именно таким путем была оценена продукция массового моноциклического рачка Calanus finmarchicus в Баренцевом (Яшнов, 1940; Камшилов, 1958) и Норвежском (Тимохина, 1968) морях, а также Diaptomus salinus – в Аральском море (Яблонская, Луконина, 1962) и некоторых других видов.
Этими авторами фактически применялся один и тот же методический подход: определялся момент достижения максимума численности популяции, а затем наблюдения проводились через короткие промежутки времени.
Однако при этом получаются заниженные величины продукции, так как она оценивается фактически для вегетационного периода, а не для всего года, поскольку расчет начинается только с момента достижения популяцией максимальной численности и не учитываются особи, родившиеся до этого (Тимохина, 1968; Грезе, 1973; Воронина и др., 1979).
Численность может оказаться заниженной также изза недостаточной частоты съемок, вследствие чего ее кратковременное нарастание до максимума может быть пропущено. Кроме того, к значительным ошибкам может приводить также и осреднение данных, полученных с довольно больших акваторий, где даже в пределах одной водной массы развитие популяции происходит не всегда синхронно (Воронина и др., 1982).
Следовательно, этот способ определения продукции трудоемок и недостаточно точен, поэтому приходится прибегать к методам, позволяющим оценивать ее на основании данных кратковременных экспериментов, а немногочисленные прямые наблюдения за скоростью роста использовать для их корректировки (Грезе, 1983). Такой подход используется для расчета продукции популяции копепод (Bergreen et al., 1988; Kiorboe, Nielsen, 1994). Перспективным представляется также метод, основанный на выделении отдельных размерных групп веслоногих раков с последующим культивированием в естественной среде (Borgman et al, 1979; Nichols, Thompson, 1991).
Помимо вышеперечисленных экспериментальных способов определения продукции водных беспозвоночных, достаточно широко применяются расчетные методы ее оценки (Винберг, 1968; Иванова, 1985; Алимов, 1989 и др.). В их основу положено балансовое равенство обменных процессов особи, популяции или сообщества:
C = (P+R)/f ,
где C – рацион, P – продукция, R – скорость метаболизма (дыхания), f – коэффициент усвояемости пищи.
Величина f для большинства организмов колеблется в пределах от 0.6 до 0.9 и для приблизительных расчетов может быть принята равной 0.8 (Conover, 1966; Винберг, 1968). В основу произведенных нами расчетов продукции, (Примаков, Бергер, 2007) были положены данные, собиравшиеся круглогодично в течение ряда лет на уже упоминавшейся неоднократно станции Д-I, расположенной в устьевой части губы Чупа. Величины рациона и скорости метаболизма исследованных планктонных ракообразных рассчи
тывали с помощью общепринятых методов (Сущеня, Хмелева, 1967; Винберг, 1968; Сущеня, 1972; 1975; Иванова, 1985; и др.). Поскольку в Белом море на долю ракообразных приходится до 85% численности и до 95% биомассы зоопланктона (Перцова, Прыгункова, 1995), мы пренебрегли вкладом в общую продукцию зоопланктона Chaetognatha, Appendicularia и Hydrozoa. Не были также учтены меропланктонные организмы и микрозоопланктон, представленный в основном инфузориями. Вклад микрозоопланктона в общую продукции планктонных животных Белого моря был оценен И.М. Примаковым позже (Бергер и др., 2007).
Результаты расчетов продукционных характеристик для основных видов зоопланктона приведены в табл. 23.
Таблица 23 Продукционные характеристики основных видов зоопланктона (с сокращениями по: Примаков, Бергер, 2007)
Типичная картина сезонной динамики продукционных характеристик зоопланктона на станции Д-1 проиллюстрирована результатами расчетов, выполненных на основании данных 1999 г. (рис. 18). Суточная продукция зоопланктонного сообщества варьировала от 0.8 до 16.4 мг/м3, а интегральная годовая продукция составила 1.6 г/.
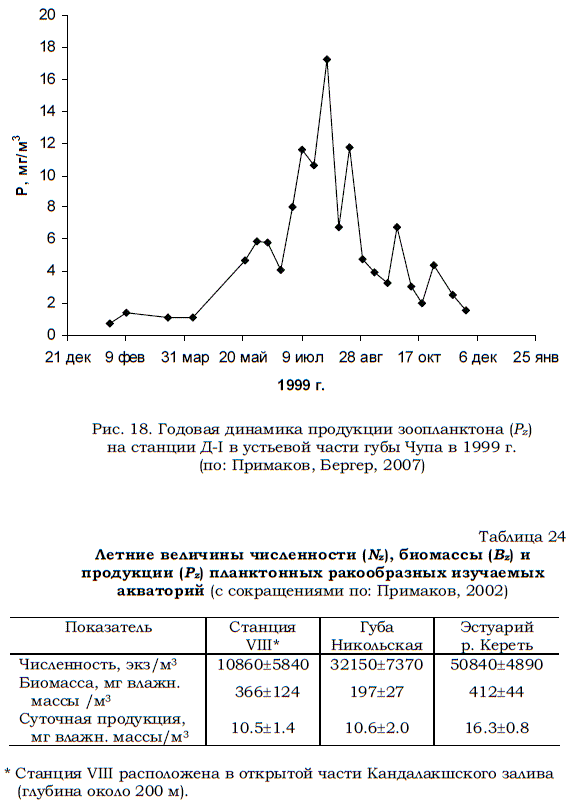 Рис. 18. Годовая динамика продукции зоопланктона (Рz) на станции Д-I в устьевой части губы Чупа в 1999 г. (по: Примаков, Бергер, 2007)
Рис. 18. Годовая динамика продукции зоопланктона (Рz) на станции Д-I в устьевой части губы Чупа в 1999 г. (по: Примаков, Бергер, 2007)
Таблица 24 Летние величины численности (Nz), биомассы (Bz) и продукции (Pz) планктонных ракообразных изучаемых акваторий (с сокращениями по: Примаков, 2002)
Аналогичные расчеты (табл. 24) продукции зоопланктона были выполнены (Примаков, 2002) на основании материалов, собранных в других частях Кандалакшского залива (эстуарии р. Кереть, губе Никольской и др.).
Просуммировав и усреднив данные обеих работ, можно заключить, что продукция зоопланктона в среднем за все сезоны составляет около 5 мг/м3 в сутки. В целом для Белого моря величина продукции составляет около 9.85 млн. т. в год, что при калорийности зоопланктонных ракообразных около 0.5 ккал/г влажной массы (см. табл. 1) эквивалентно 4.9.1012 ккал/год.
Валовую продукцию зоопланктона, можно рассчитать и иным способом, через годовой P/B-коэффициент. Ранее (Бергер и др., 1995; Бергер, 2005) мы исходили из того, что, как сказано в учебнике по гидробиологии А.С. Константинова (1986), этот коэффициент для зоопланктона равен 2.5. Последующее знакомство с дополнительной литературой и устные сведения, полученные при обсуждении от ряда специалистов, показали, что эта величина занижена в 3–4 раза. Величина P/B-коэффициента существенно варьирует у одного или того же вида планктонных ракообразных в зависимости от широты и соответственно температуры воды мест обитания. В Центральной Арктике годовой P/Bкоэффициент Calanus hyperboreus был равен 0.3 – 0.76 (Кособокова, 1986). По данным того же автора в Белом море его величина у С. glacialis составляла 5.4, а у C. finmarchicus в Норвежском море она варьировала от 0.6 до 8.8.
Для зоопланктона мелководья Баренцева моря приведена величина годового P/B-коэффициента, равная 6.9 (Камшилов, 1958). Годовой P/B–коэффициент для зоопланктона Мирового океана по данным П.А. Моисеева (1989) в среднем равен 10. Сходный данные приводятся (Шунтов, 1985) для зоопланктона Охотского моря, более близкого к Белому морю по термическому режиму, чем океан в целом.
Не останавливаясь больше на анализе этих данных, отметим, что и в наших последних расчетах годовой P/Bкоэффициент был для различных исследованных видов равен в среднем 9.1 (табл. 23)*. Приняв эту величину за основу, нетрудно подсчитать годовую продукцию всего беломорского зоопланктона, исходя из данных о его средней биомассе. Общее содержание зоопланктона в Белом море при средней биомассе около 200 мг/м3 составляет 1.08 млн. т. Следовательно, масса всего зоопланктона, содержащегося в Белом море, эквивалентна 0.52.1012 ккал, а продукция зоопланктона составляет порядка 4.7.1012 ккал/год.
Таким образом, расчеты годовой продукции зоопланктона, выполненные двумя способами, дали весьма близкие результаты, усреднив которые получим величину 0.48 . 1013 ккал.
Исходя из установленных соотношений (табл. 23) и уровня продукции зоопланктона, можно определить годовой рацион всех планктонных ракообразных Белого моря, который равен 2.2 . 1013 ккал.
Годовая продукция зоопланктона всего моря эквивалентна 4.8.1012 ккал, что в пересчете на 1 м2 составляет около 50 ккал. Годовой рацион зоопланктона в энергетическом выражении соответствует 2.2.1013 ккал или примерно 245 ккал/м2.
При этом следует отметить, что точность расчетов рациона и продукции зоопланктона весьма приблизительна
____________________ * В качестве комментария к этому выводу следует заметить, что принятая в наших расчетах величина калорийности планктонных Copepoda, равная 0.5 ккал/г влажной массы (см гл.1), по-видимому, занижена. С помощью метода бихроматного окисления показано (Кособокова, 1980), что калорийность беломорских планктонных ракообразных Сalanus glacialis, Metridia longa и Pseudocalanus elongatus равна соответственно 1.58, 0.75 и 0.67 ккал/г влажной массы, а их средняя калорийность – 1 ккал/г влажной массы, что в 2 раза превосходит величины калорийности планктонных копепод, использованные в наших расчетах. Эти данные не были приняты во внимание по той причине, что они значительно отличаются от аналогичных величин, приводимых другими авторами (см. табл.1) и нуждаются в проверке. При этом не исключено, что они ближе к истине и отражают особенность не только беломорских копепод, заключающуюся в более высоком содержании жира в их теле (Кособокова, 1980), но и, возможно, полярного рачкового планктона в целом. Последнее предположение подтверждают данные о том, что калорийность антарктических Euphasia superba равна 1.1 ккал/г влажной массы (Чекунова, Рыкова, 1974).
по причине того, что соотношение хищных и нехищных видов остается неизвестным. К этому необходимо также добавить, что данные о биомассе и численности зоопланктона скорее всего значительно занижены из-за недоучета малых форм: так называемого пикопланктона, обычно проходящего через мельничное сито, используемое в стандартных орудиях лова зоопланктона. Сказанное относится и к фитопланктону (Сапожников, 2005), однако в случае с зоопланктоном без дополнительных данных преждевременно вносить соответствующие поправки в расчеты.
Частично заполнить этот пробел позволяют данные о продукции планктонных инфузорий, полученные И.М. Примаковым (Бергер и др., 2007). Согласно их результатам продукция микрозоопланктона в Белом море была приблизительно оценена в 550 мг/м3 за год. Исходя из этого, можно подсчитать, что годовая продукция микрозоопланктона всего моря эквивалентна энергетически 1.5.1012 ккал.
Расчеты рациона микрозоопланктона ранее не производились из-за недостатка необходимых сведений. Однако, исходя из того, что у растительноядных форм коэффициенты усвояемости пищи и эффективности продукции равны соответственно 0.6 и 0.20–0.24 (Алимов, 1989), можно приблизительно определить рацион микрозоопланктона. Расчеты, произведенные И.М. Примаковым, дали величину рациона, равную 1.1013 ккал/год.
Таким образом, суммарная величина годовой продукции беломорского мезои микрозоопланктона составляет около 6.3.1012 ккал, а рацион – 3.2.1013 ккал.
Согласно представленным выше результатам, микрои мезозоопланктоном за год суммарно потребляется весьма существенная (не менее половины) часть энергии, продуцируемой фитопланктоном. На первый взгляд такие трофические отношения могут показаться слишком напряженными для нормального функционирования экосистем беломорской пелагиали, однако рассчитывать количественные взаимоотношения между фитои зоопланктоном путем деления энергетического эквивалента продукции фитопланктона на величину потребления энергии зоопланктоном нельзя, поскольку планктонные микроводоросли – не единственный источник питания зоопланктона. Как
109
уже отмечалось, существенную роль в питании последнего составляют бактерии, находящиеся в толще воды в свободном состоянии либо ассоциированные с детритом (Сорокин, 1973 б; Парсонс и др., 1982; Теплинская, 1995 и др.). Таким образом, есть все основания считать, что суммарная продукция бактериои фитопланктона, составляющая в Белом море около 3.0.1014 ккал/год, примерно на порядок превышает потребление энергии зоопланктоном. Если к этому добавить продукцию криофлоры, составляющую около 1/4 от продукции фитопланктона (Бергер, 2006), то сумма составит около 3.2.1014 ккал/год.
Итак, оказывается, что потребление энергии и вещества мезои микрозоопланктоном обеспечено количеством пищи, на порядок превосходящим его. Следовательно, трофические отношения между беломорскими консументами первого порядка, составляющими основу зоопланктона, с одной стороны, и бактериои фитопланктоном, включая криофлору, с другой стороны, вполне нормальные, а не напряженные.
К этому следует добавить, что из-за отсутствия данных о том, какую долю составляют хищные формы зоопланктона, произведенные расчеты весьма приблизительны, поскольку совершенно не учтено потребление пищи внутри сообщества зоопланктонных организмов. Очевидно, что чем больше доля хищников, тем меньше, по сравнению с указанным выше, будет общее потребление пищи зоопланктоном, который выедается не только рыбами, гребневиками, медузами и другими животными, но и собственными зоопланктонными хищниками.



