Сельдь
Беломорская сельдь Clupea pallasi maris-albi Berg относится к числу важнейших промысловых рыб Белого моря. Долгие годы вопрос о ее внутривидовой структуре был предметом дискуссии. К настоящему времени на основании результатов различных исследований
(Рабинерсон, 1925; Дмитриев, 1946; Тамбовцев, 1957; Артемьева, 1975; Душкина, 1975; Мухомедьяров, 1975a, б; Лайус, 1990, 2002; Семенова, 2004 и др.) можно считать установленным, что беломорская малопозвонковая сельдь представляет собой комплекс изолированных, но в то же время близких форм, связанных общностью происхождения от тихоокеанских предков. Количество таких форм остается пока недостаточно определенным, но факт их существования можно считать установленным.
Сельдь осуществляет значительные сезонные перемещения по акватории Белого моря (Мухомедьяров, 1975б; Ермольчев, Похилюк, 1985). Локальные стада приурочены к определенным гидрологическим условиям и местам нереста в различных районах моря. Их миграции связаны главным образом с распределением кормовых объектов в период нагула. Летом основным местом откорма беломорской сельди является Бассейн. Зимовальные миграции осуществляются по двум основным путям (Зеленков и др., 1995). Сельдь, нагуливающаяся в основном в бассейновой части моря, у Терского берега, на стыке Горла и Двинского залива и, по-видимому, севернее Соловецких островов, мигрирует на зимовку в кутовую часть Кандалакшского залива и губу Чупа. Второй путь миграции – в восточную Соловецкую Салму. Эти миграции происходят в октябре. В ноябре сельдь собирается в Сорокской губе, являющейся основным местом зимовки. Часть сельдей зимует в Кандалакшском заливе, в том числе в губе Чупа. В феврале–марте сельдь покидает места зимовки, перемещаясь ближе к нерестилищам.
В отдельные годы в Белое море заходит довольно много неполовозрелой атлантической сельди из Баренцева моря (Тамбовцев, 1956, 1966), и, наоборот, беломорская сельдь проникает в юго-восточные районы Баренцева моря (Стасенкова, 2005).
Половая зрелость беломорской сельди наступает на втором–третьем (онежские, двинские и мезенские сельди) или на третьем–четвертом годах жизни, как у сельдей Кандалакшского залива (Мухомедьяров, 1975 а). Основу нерестовых стад составляют рыбы в возрасте 3–4 лет, хотя в отдельные годы значительная доля нерестящихся сельдей может приходиться и на старшие (5–6 лет) возрастные группы. Плодовитость сельди варьирует от 12 до 62 тыс. икринок (Марти, 1952; Кузнецов, 1960).
Значительная часть беломорских сельдей нерестится весной. Основной нерестовый субстрат – морская трава Zostera marina. После ее массовой гибели в 1960 г. сельдь стала откладывать икру на литоральные и сублиторальные бурые водоросли, главным образом фукоиды.
В отдельных заливах нерест сельди происходит в разное время. В Кандалакшском заливе первыми в апреле–мае, еще подо льдом или в период его таяния при температуре воды около 0oС, нерестятся мелкие (12–20 см в длину) «егорьевские» сельди (Мухомедьяров, 1975 а). «Ивановская» сельдь, имеющая длину тела от 20 до 30 см, обычно менее многочисленна. Ее нерест происходит в июне при температуре воды 5–12o С. Сельди Онежского залива нерестятся в мае–июне после того, как залив очистится от льда и температура воды превысит 4–5oС. В Двинском заливе нерест сельди происходит в мае–июне при температуре воды не ниже 7–9oС. В Мезенском заливе икрометание происходит в мае–июне при температуре воды от 4 до 12oС (Кузнецов, 1960).
В Кандалакшском заливе основу пищи личинок сельди при переходе на активное питание составляют личинки Pseudocalanus minutus и других Copepoda, в Онежском заливе – личинки моллюсков и инфузории (Чуксина, 1970; Гошева, Слонова, 1975; Пущаева, 1981). Сеголетки сельди питаются исключительно Copepoda, главным образом Acartia longiremis и A. bifilosa (Эпштейн, 1957). Для взрослой сельди Кандалакшского и Двинского заливов в пищевом рационе наиболее характерны эуфазииды (Тарвердиева и др., 2000) и веслоногие раки Calanus glacialis, Acartia longiremis, Metridia longa и Pseudocalanus minutus. Основу питания взрослой сельди в Онежском заливе составляют нектобентические мизиды и бокоплавы. В Мезенском заливе сельдь потребляет преимущественно личинок усоногих раков (Слонова, 1977).
Сельдь потребляется хищными рыбами (в основном треской) и некоторыми морскими млекопитающими (в основном кольчатой нерпой и белухой) и рыбоядными птицами (бакланами). Личинки и мальки сельди служат объектом питания для гораздо большего числа беломорских животных, чем взрослая сельдь. Основными хищниками по отношению к ним являются треска, навага, корюшка и некоторые другие рыбы, в том числе и взрослая сельдь. Потребляет молодь сельди полярная крачка. Среди беспозвоночных животных, поедающих и (или) убивающих личинок и мальков сельди, одно из первых мест занимают различные кишечнополостные (Иванченко, 1995).
Беломорскую сельдь начали промышлять еще в XV веке (Кузнецов, 1960). В первое время лов производился лишь в районе Соловецких островов, а позднее – и в других частях моря. В 1830–1840-х гг. вылов сельди достигал 34 тыс. т в год. Из тех сведений, которые имеются в литературе, следует, что это – максимальный уровень сельдяного промысла на Белом море. Возможно, что столь значительные уловы были отчасти обусловлены массовыми заходами в Белое море молоди атлантической сельди. Не исключено также, что добыча сельди могла достигать и больших величин, но данных, подтверждающих или опровергающих это предположение, нет. Спустя всего 30 лет после названного выше срока, вылов сократился в 3 раза (Богуслав, 1846; Сидоров, 1879; Кузнецов, 1960). В первой половине ХХ века объем годового промысла сельди понизился до 3–5, а после 1960 г. он уменьшился до 1–2 тыс. т. В наше время ежегодно вылавливается менее 1 тыс. т (Зеленков, Стасенков, 1997; Отчет СевПИНРО, 2003). Таким образом, по сравнению с максимальным уровнем, зарегистрированным около 170 лет назад, ежегодная добыча сельди уменьшилась в наше время в 50–100 раз.
Причины падения вылова сельди в Белом море неоднократно анализировались в научной литературе (Кузнецов, 1960, 1961; Зеленков и др., 1995; Berger et all., 2001; Житний, 2005 б, 2007 и др.), и рассматривать их еще раз нет необходимости. Достаточно заметить, что наблюдающаяся почти двухвековая депрессия промысла обусловлена переловом и различными нарушениями воспроизводства сельди, носящими как антропогенный, так и естественный характер.
В период расцвета промысла проблема сохранения запаса беломорской сельди никого не беспокоила. Об этом стали задумываться значительно позже: во второй половине прошлого столетия, когда уловы сельди резко сократились.
Основные оценки запаса производили на основе данных промысловой статистики уловов (Зеленков и др., 1995; Зеленков, Стасенков, 1997; Стасенков, 1998), а также с помощью гидроакустических съемок, результаты которых верифицировали путем контрольных тралений (Ермольчев, Похилюк, 1985; Ермольчев и др., 1987; Ермольчев и др., 1990). Во второй половине 70-х годов прошлого столетия запас сельди в Белом море был крайне низок: 1.8 тыс. т. За прошедший примерно 30–летний период он увеличился более, чем в 6 раз, достигнув к 2003 г. 12 тыс. т (рис. 20). Эта величина будет использована в дальнейшем при расчете продукции беломорских рыб.
Можно предполагать, что рост запаса сельди обусловлен главным образом постепенным восстановлением беломорской зостеры (Вехов, 1995), гибель которой произошла в 1960 г. Заросли этой травы служат местом нереста и инкубации икры, а также местом откорма и убежищем для личинок и мальков сельди. Одновременно с увеличением запаса сельди в течение нескольких последних лет наблюдается вспышка численности трехиглой колюшки, которая делает нерестовые гнезда в зарослях зостеры (Иванова и др., 2007).
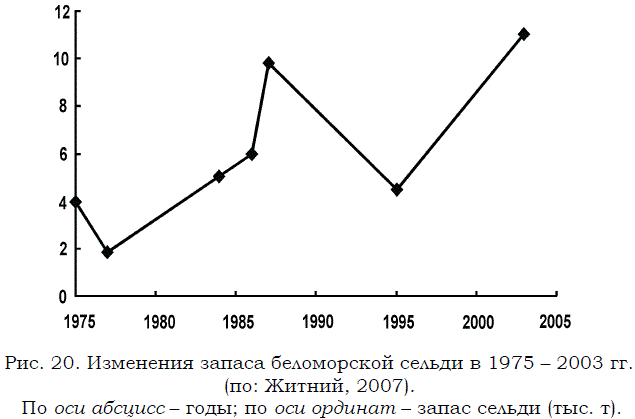
Рис. 20. Изменения запаса беломорской сельди в 1975 – 2003 гг. (по: Житний, 2007). По оси абсцисс – годы; по оси ординат – запас сельди (тыс. т).



