Зоопланктон в центральной части моря — Бассейне и открытых районах заливов
Впервые данные о количественном распределении зоопланктона получены весной 1926 г. В. А. Яшновым (1940) для Бассейна, Кандалакшского, Двинского и Онежского заливов. После боль. шого перерыва эта акватория моря была вновь охвачена съем. ками — летом 1959 (Эпштейн, 1963), 1972 (Перцова, 1980б) и 1981 гг. (Прыгункова и др., 1985). Средняя биомасса зоопланк. тона, рассчитанная для каждого из обследованных районов за весь упомянутый период, приведена в табл. 6.5 и 6.6. Сравнение величин биомассы показывает, что в районах с «бассейновым» типом структуры вод, отличающихся в летний период резко выра. женной стратификацией температуры и солености (Тимонов, 1950), зоопланктон богаче. Это районы, свободно сообщающиеся между собой: Бассейн, Кандалакшский и Двинский заливы. Биомасса зоопланктона значительно ниже в районах с перемешанным типом вод (Тимонов, 1950), характеризующихся однообразным, а иногда и совершенно однородным вертикальным распределением температуры, солености и других гидрологических елементов. Это районы, отделенные от центральной части моря онтальными зонами (Кравец, 1985) — Онежский залив и Горло, таким же районам следует отнести Мезенский залив и Воронку, наименьшая средняя биомассахарактерна для Мезенского залива см. табл. 6.5 и 6,6), что подтверждают исследования последних лет
(Антипова и др., 1985: Слонова, 1985).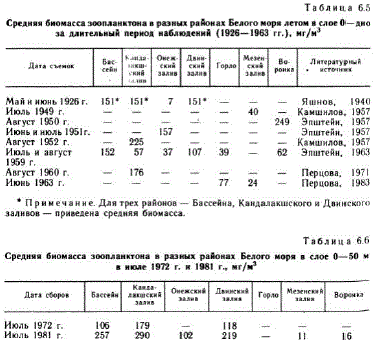
Горизонтальное распределение биомассы зоопланктона в Белом море летом неравномерное (Хмызникова, 1947; Перцова, 1970, 1971, 1980б; Прыгункова, 1985а). Согласно данным съемки, проведенной в 1972 г. (Перцова, 1980б) в центральной части моря биомасса минимальна (рис. 6.12). Она увеличивается по мере приближения квершинам Кандалакшского и Двинского заливов и по направлению к берегам. Пятна с высокой биомассой располагаются в районах сезонных термических фронтов, формирующихся (Кравец, 1985) на границе зон, в одной из которых приливные течения перемешивают воды отповерхности до дна, а в другом нет. Такие районы расположены в южной оконечности Горла и награнице Онежского залива с Бассейном севернее Соловецких островов. Летом 1972 г. наиболее богатыми районами были мелкоBOLEH
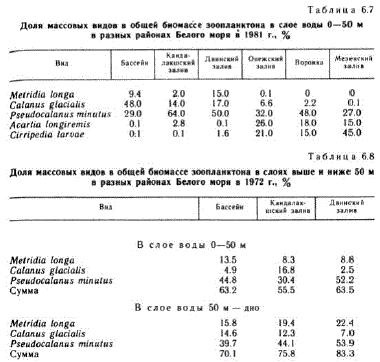
В распределении по вертикали общим для всех упомянутых районов является то, что наибольшая биомасса везде отмечается в верхнем слое до 25—50 м. С глубиной биомасса уменьшается. Биомасса зоопланктона в Белом море складывается в основном за счет немногих массовых видов, среди которых главную роль играют копеподы. Из них чаще других во всех районах на первое место выходит Pseudocalaлия тілигия (табл. 6.7 и 68). Летом в центральной части моря и глубоководных частях Кандалакшского и Двинского заливов доминируют Р. тіпи?иs, Calапиs glacialis, МеIridia Іопgа. Доля остальных копепод невелика. В мелководных частях тех же заливов, а также в других мелководных районах — Онежском заливе, Воронке, Мезенском заЛиве — возрастает роль мелких копепод, например Acartia longireтis, а во второй половине лета и Temora longicornis (Эпштейн, 1957а). В Мезенском заливе очень большая доля в лет. ний период приходится наличинки усоногих раков. В Горле Белого Моря обитают в основном массовые виды, являющиеся общими для Белого и Баренцева морей (Перцова, 1983). У Терского берега, где влияние питающего течения больше, до 10—20% 'ocTaB,11 kot P. minutus H C. glacilis. 31ecb o4eHb Ben HKa (54 %) я щетинкочелюстных — Sagitta elegans. У Зимнего берега, области сточного течения, зоопланктон на 45-55 % состоит псевдокалянуса. Кроме того, здесь сильно возрастает роль чинок донных животных. Их количество у Зимнего берега в раз выше, чем у Терского. Биомасса зоопланктона в Горле ше в районе Терского берега (116—170 мг/м"), а в районе ЗимЕго берега она в несколько раз ниже (15—27 мг/м").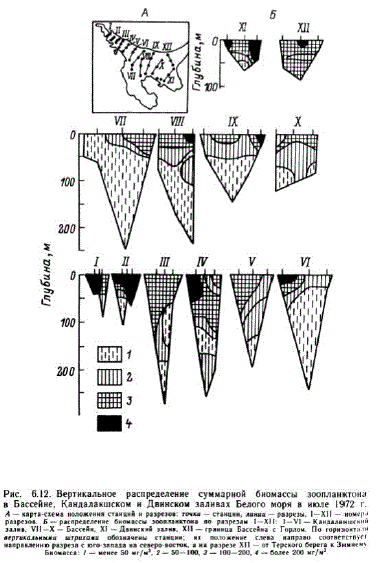
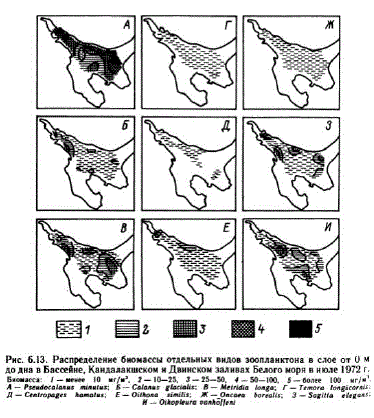
В июле 1972 г. в центральной части моря, Кандалакшском и Двинском заливах распределение трех холодноводных видов — P. тіпи!иs, С. glacialis и M. Iопgа (рис. 6.3) — отражало распределение всей биомассы зоопланктона, поскольку их доля в его общей биомассе выше, чем всех остальных видов (см. табл. 6.8). Для этих видов характерно увеличение биомассы в области фронтальных зон — на границе Бассейна с Горлом и с Онежским заливом. Минимальная биомасса у всех видов наблюдается в центральной части моря, совпадаюшей с областью антициклонического круговорота. Максимум биомассы у разных видов находится в разных районах: у Р. тіпития награнице Бассейна с Горлом, у С. glacialis в Кандалакшском заливе, пятна максимальной биомассы M. longa отмечаются в четырех районах: в восточной части Бассейна, совпадающей с областью циклонического круговорота («полюса холода»), на границе Бассейна с Онежским заливом, в районе Терского берега и во внешней части КандалакшcKoro 3a.JIHBa y Kape.IbcKoro 6epera. B oT.1 Huq He oT P. minutus H С. glacialis во внутренней части Кандалакшского залива и на границе Горла с Бассейном отмечаются зоны минимальной биоMaccl, M. longa (pHc. 6. 13).
Остальные виды копепод, включая представителей подотряда Cyclopoidа, составляют не более 7 % общей биомассы в этот период. Распределены они в исследуемом районе довольно равно, мерно (см. рис. 6.13) и поэтому существенно не меняют картину распределения общей биомассы. Кроме приведенных в табл. 68 массовых видов, доминирующих в июле, в указанных районах значительная роль в общей биомассе принадлежит также Sagitta elegars H Oikopleura van hoffeni. Ha oTue.TbH bx cTa HLL HHX oH H H сумме составляют до 11—24 % общей биомассы. Распределение сагитты сходно с распределением массовых холодноводных видов копепод. Максимальная биомасса ойкоплевры отмечается во фронтальных зонах и на границе Кандалакшского залива с Бассейном.