Бактериопланктон
Микроорганизмы, обитающие на дне водоемов (бактериобентос) и в толще воды (бактериопланктон) – важный структурно-функциональный элемент экосистем моря. Они играют одну из основных ролей в балансе вещества и энергии, с одной стороны, как трансформаторы органики, а с другой стороны, как ее продуценты и источники пищи для различных обитателей водоема.
Несмотря на то, что роль микроорганизмов в трофике морских экосистем трудно переоценить, до сих пор она изучена далеко недостаточно. Кроме того, к сожалению, подавляющая часть соответствующих исследований выполнена в морях тропической и умеренной зон Мирового океана. Из отечественных морей больше всего повезло в этом отношении Черному морю (Сорокин, 1982; Гутбейн, 1990).
Арктические моря исследованы гораздо хуже. Плохая изученность арктических микроорганизмов связана не только с трудностями работ в высоких широтах. Она обусловлена и иной причиной: долгое время существовавшей ошибочностью наших представлений об этой составляющей биоты океанов и морей. Как и у других пойкилотермных организмов (термокомформеров), функциональная активность бактерий напрямую связана с воздействием ряда внешних факторов, в первую очередь температуры. Поскольку арктические водоемы характеризуются крайне низкими температурами воды, то в соответствии с законом Вант-Гоффа длительное время a priori предполагалось, что роль микроорганизмов в трансформации вещества в них ничтожна по сравнению с таковой в морях умеренных широт, однако, как оказалось, это не соответствует действительности.
Было установлено, что в водах арктических морей содержится множество бактерий, названных психрофильными, которые способны размножаться при низких температурах, близких к точке замерзания воды (ZoBell, 1946; Крисс, 1959, 1976; Теплинская, Москвина, 1987 и др.). Накоплено уже довольно много сведений об этих микроорганизмах, обитающих в Мировом океане при температурах 5oС и ниже (Morita, 1975), но специфика их жизни при низких температурах только начинает проясняться (Теплинская, Москвина, 1987).
Очень мало известно, в частности, о распределении и активности микроорганизмов в отдельных морях, включая арктические. Не составляет исключения и Белое море. Имеющиеся сведения (Галкина, 1985; Теплинская, Москвина, 1987; Галкина и др., 1988; Теплинская, 1995) позволяют лишь в самых общих чертах охарактеризовать роль микроорганизмов в функционировании беломорских экосистем Первые сведения о беломорском бактериопланктоне получены при исследовании протеолитической активности бактерий, обеспечивающих самоочищение воды (Пилипас, 1977). Данные о плотности, биомассе и продукции бактериопланктона начали накапливаться лишь с 1982–1983 гг., когда было предпринято исследование потоков органических веществ, как в местах культивирования мидий в прибрежных водах, так и в свободных от марикультуры открытых частях Кандалакшского залива (Галкина, 1985). Чуть позже развитие летне-осеннего бактериопланктона было изучено в проливе Великая Салма (Теплинская, Москвина, 1987). В дальнейшем бактериопланктон исследовали и в других районах Кандалакшского залива в разные сезоны, включая зиму и раннюю весну, когда пробы отбирали подо льдом (Галкина и др., 1982, 1988; Галкина, Кулаковский, 1993).
Методы исследования, использованные в этих работах, были одинаковы, несмотря на различные задачи, что позволяет сопоставлять результаты разных авторов. Во всех случаях численность бактериопланктона подсчитывали на мембранных фильтрах после фильтрации 1–15 мл воды (Разумов, 1962). Биомассу бактерий рассчитывали по стандартной методике, исходя из общего числа клеток и их среднего объема (Романенко, Кузнецов, 1974). Бактериальную продукцию определяли прямым скляночным методом в радиоуглеродной модификации либо рассчитывали по результатам биологического потребления кислорода (Руководство..., 1980). В дальнейшем биомасса и продукция планктонных бактерий выражались как в единицах сырой массы, так и в мг углерода, исходя из того, что на долю органического углерода приходится 10% от сырой массы бактерий (Сорокин, 1973 б).
Результаты выполненных исследований показали, что в Белом море (Кандалакшский залив) размеры бактериальных клеток варьируют в довольно значительных пределах. В губе Чупа их объем равен 0.18–0.26 мкм3 (Галкина, 1985). В проливе Великая Салма были обнаружены бактериальные клетки гораздо большего объема, достигавшего 0.62 мкм3 (Теплинская, Москвина, 1987).
Что касается данных о сезонных изменениях обилия бактериального планктона, то работами, выполненными в Кандалакшском заливе, было показано (Теплинская, 1995), что в летний период плотность бактерий в толще воды составляла в среднем около 0.9–1.2 млн. клеток в 1 мл. При этом наибольшие концентрации бактерий были зарегист
рированы в приповерхностном (150 мкм) и придонном слоях воды. Максимальное количество бактерий наблюдалось в прибрежных районах моря, а по мере продвижения в открытую часть залива их численность уменьшалась.
Осенью общая плотность бактериопланктона в Кандалакшском заливе варьирует от сотен тысяч до миллионов клеток в 1 мл воды. Она достигает максимума в приповерхностном слое воды и уменьшается по направлению от прибрежных участков к открытым районам залива. Основным фактором, влияющим на обилие бактериопланктона в осенний период, является смена водных масс во время приливов и отливов. Корреляция с изменениями солености и температуры отсутствует (Теплинская, 1995). Изменения численности и биомассы с глубиной показывают, что максимальное развитие бактериопланктона приходится на 2-метровый поверхностный горизонт, характеризующийся максимальным развитием фитопланктона (Ильяш и др., 2003), что свидетельствует о связях развития этих компонентов планктонного сообщества. По-видимому, отмершие клетки фитопланктона, образующие детрит, служат субстратом для массового развития бактерий.
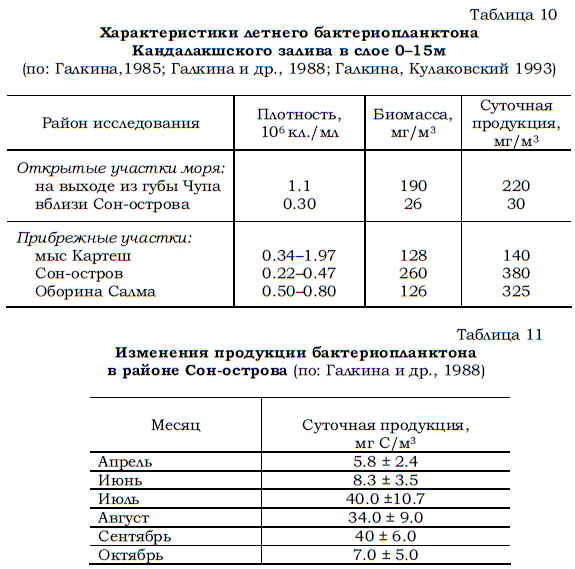
Прибрежные районы отличаются от открытых частей моря не только большей плотностью планктонных бактерий, но и более высокими показателями их биомассы и продукции. Данные, подтверждающие этот вывод, были получены в различных частях Кандалакшского залива Белого моря (табл. 10).
Наиболее полно сезонные изменения продукции бактериопланктона были исследованы с апреля по октябрь 1985–1986 гг. в районе Сон-острова (Кандалакшский залив). Показано (Галкина и др., 1988), что биомасса, продуцируемая планктонными бактериями, была максимальной (340– 400 мг/м3 в сутки) в июле–сентябре, однако и в другие сезоны она не опускалась ниже 58–70 мг/м3 в сутки. После пересчетов, произведенных на основании указанного выше соотношения (Сорокин, 1973), оказалось, что суточная продукция планктонных бактерий варьировала в течение исследованного периода от 5.8 до 40.0 мг С /м3 (табл. 11).
Таблица 10 Характеристики летнего бактериопланктона Кандалакшского залива в слое 0–15м (по: Галкина,1985; Галкина и др., 1988; Галкина, Кулаковский 1993)
Таблица 11 Изменения продукции бактериопланктона в районе Сон-острова (по: Галкина и др., 1988)
Исходя из данных, полученных радиоуглеродным методом, была рассчитана средняя величина годовой продукции бактерий в прибрежных и открытых участков трех районов Кандалакшского залива Белого моря (губа Чупа, пролив Оборина Салма и акватория у Сон-острова). Она оказалась равной 4.5 г С/м3, что эквивалентно* 45 ккал/м3.
____________________ * При переводе величин продукции в энергетический эквивалент используются результаты расчетов балансового уравнения фотосинтеза, из которых следует, что 1 мг углерода, содержащегося в синтезированных органических веществах, соответствует примерно 10 кал (Винберг, 1960).
Величина продукции бактерий, определенная с помощью кислородного метода, оказалась значительно выше (Галкина и др., 1993). Исходя из этих данных, была рассчитана величина годовой продукции бактериопланктона (Бергер и др., 2007). При этом следует оговорить, что имеющиеся материалы касаются только верхнего 15метрового слоя воды, годовая продукция бактериопланктона в котором энергетически эквивалентна 0.6. 1014 ккал.
Исходя из того, что средняя глубина Белого моря равна 60 м (Бабков, Голиков, 1983), нетрудно подсчитать, что годовая продукция бактериопланктона всей толщи воды составляет 2.4 . 1014 ккал.
Однако до тех пор, пока не будет получено достаточно данных об обилии и продукции планктонных бактерий во всей толще вод Белого моря, такой расчет может считаться лишь весьма приблизительным. Скорее всего, указанная выше величина продукции бактериопланктона значительно завышена по сравнению с реальным уровнем.
Высокие темпы размножения и значительная продукция бактериопланктона свидетельствуют о том, что бактериальное сообщество активно функционирует в Белом море в течение всего года. Сравнение с аналогичными данными для Баренцева моря (Teplinskaya, Dudarenko, 1984; Байтаз, 1990) показывает, что суточная продукция бактериальной биомассы в Кандалакшском заливе в 2–3 раза выше таковой в Баренцевом море. Показатели плотности бактериопланктона в Белом море сопоставимы с таковыми и в других морях, например в северо-западной части Черного моря (Нижегородова и др., 1981). При этом биомассы бактерий Кандалакшского залива в несколько раз превышают таковые в Черном море. При близких численностях это обусловлено различиями размеров бактериальных клеток, объем которых в Кандалакшском заливе в среднем выше, чем в Черном море (Теплинская, 1995). Что касается скорости роста биомассы, то показатели беломорских планктонных бактерий близки бактериопланктону в морях умеренных широт, в частности в прибрежных районах Черного моря (Сорокин, 1982; Гутбейн, 1990).
Приведенные выше данные о продуктивности планктонных бактерий позволяют перейти к обсуждению их места в балансе вещества и энергии в экосистемах пелагиали Белого моря, однако перед этим необходимо сделать ряд замечаний общего характера. Дело в том, что, как уже отмечалось, водные бактерии длительное время считались лишь деструкторами органического вещества. Эта сторона их жизнедеятельности была достаточно полно исследована уже к середине ХХ столетия (Харвей, 1948). Многочисленными наблюдениями было показано, что за счет деструкции органики микрофлорой происходит трансформация большей части энергии, поступающей в водные экосистемы с автохтонными и аллохтонными органическими веществами. Несмотря на успехи этих исследований, морские бактерии еще долгое время оставались плохо изученными, а их продукция в Мировом океане продолжала считаться гораздо менее значимой, чем продукция планктонных микроводорослей. Примером этого могут служить устаревшие данные, представляющие в настоящее время лишь историческую ценность (табл. 12).
Таблица 12 Биомасса и продукция в океане (с изменениями по: Виноградов, 1967)

Постепенно, однако, накапливалось все больше данных, согласно которым около трети органического вещества, подвергающегося в водоеме бактериальной деструкции, идет на биосинтез микробной массы (Seki, ZoBell, 1967; Riley, 1970; Сорокин, 1973б и др.). Кроме того, различными методами (в первую очередь с помощью меток по 14С) была показана важная роль микроорганизмов в формировании основных пищевых ресурсов в различных водоемах (Seki, 1966: Winberg, 1972; Сорокин, 1973 б; Парсонс и др., 1982; Гутбейн, 1990; Теплинская, 1995 и др.). Оказалось (Сорокин, 1977), что бактериальная продукция особенно важна для питания зоопланктона, поскольку бактериопланктон – основной источник питания гетеротрофного жгутикового пикопланктона (< 1 мкм). Часть бактериальной продукции, выедаемая зоофлагеллятами и в дальнейшем включающаяся в пищевую цепь, весьма значительна.
Бактериопланктон служит также важной составляющей пищи не только планктонных, но и многих бентосных животных: губок, гидромедуз, гидроидов, аппендикулярий, полихет, двустворчатых моллюсков, в том числе мидий, асцидий и даже личинок рыб (Михеев, Сорокин, 1966; Zelikman et al., 1969; Пономарева и др., 1971; Monakov, Sorokin, 1972; Сорокин, 1973б, 1982; Bernard, 1983; Сорокин, Мамаева, 1984 и др.). Концентрации бактериопланктона, оптимальные для питания этих животных, колеблются в пределах от 0.5–2 г сырой биомассы на 1м3 у тонких фильтраторов (ветвистоусых рачков, губок, сабеллид, аппендикулярий) до 3–4 г/м3 у грубых фильтраторов, способных отфильтровывать бактерии преимущественно в скоплениях (агрегатах). К ним относятся асцидии, копеподы, эуфаузииды, гидроиды и моллюски. Такие концентрации бактериопланктона обычны для мезотрофных и эвтрофных водоемов, а в олиготрофных районах океана встречаются в зонах максимума плотности бактериопланктона (Сорокин, 1973 б).
В целом в различных морях на долю бактериопланктона в экосистемах пелагиали приходится от 60 до 90% от общего потока энергии в гетеротрофной части планктонного сообщества (Сорокин, 1973б, 1982; Копылов и др., 1981; Теплинская, 1995). Что касается общей продукции планктонных бактерий, ранее считавшейся незначительной (см. табл. 12), то в последнее время (в основном благодаря совершенствованию методов исследования) было показано, что она гораздо больше предполагавшейся раньше. Во многих морских водоемах продукция планктонных бактерий сопоставима с продукцией фитопланктона или даже превосходит ее (Парсонс и др., 1982; Сорокин, 1982; Галкина и др., 1988; Pomeroy, Wiebe, 1988; Fuhram, 1992; Теплинская, 1995 и др.). При этом, однако, нельзя не учитывать, что часть бактериальной продукции, близкая по величине к потреблению энергии простейшими, изымается из оборота вирусами (Fuhram, Noble, 1995).
Вклад бактериопланктона в продуцирование и переработку органического вещества может быть особенно значимым в периоды понижения активности фитопланктона, а также на глубинах (в афотической зоне), куда не проникает свет и где нет массового развития планктонных микроводорослей. Не менее важна роль бактерий, в том числе и планктонных, в процессах гетеротрофной переработки детрита, образующегося при разрушении слоевищ макрофитов, отмирании клеток планктона и других организмов. Детритные образования, покрытые и перерабатываемые бактериями, вовлекаются в общую пищевую цепь и становятся источником энергии для различных детритофагов, весьма многочисленных в Белом море как среди планктонных, так и среди бентосных животных. Бактерии, как отмечалось выше, играют существенную роль в эстуариях в зоне действия маргинальных фильтров, где хлопья «эстуарного снега», образующиеся в результате флоккуляции, подвергаются бактериальной переработке (Мицкевич, Намсараев, 1994) и становятся источником пищи для зоопланктонных фильтраторов (Виноградов и др., 1994).
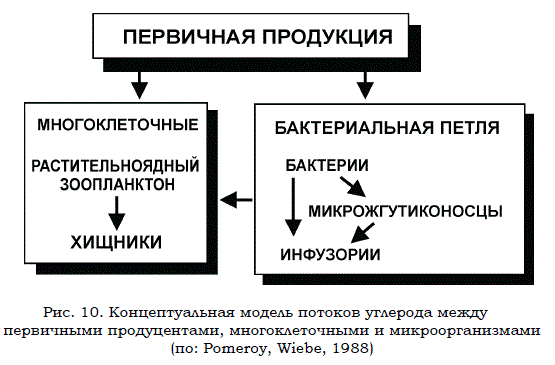
Рис. 10. Концептуальная модель потоков углерода между первичными продуцентами, многоклеточными и микроорганизмами (по: Pomeroy, Wiebе, 1988)
Таким образом, резюмируя, следует подчеркнуть, что бактериопланктон занимает важное место в трофической сети водоемов. Взаимоотношения бактериопланктона с другими элементами пелагиали моря и так называемая «бактериальная петля» изображены на рис. 10. Приведенная схема отражает связи в экосистемах пелагиали лишь частично. К ней следовало бы добавить органические вещества и тех многочисленных консументов, кроме жгутиконосцев и инфузорий, которые, как отмечалось выше, потребляют планктонных бактерий в виде пищи.
Переходя от общего к частному, т.е. к обсуждению роли бактериопланктона в функционировании пелагических экосистем Белого моря, необходимо признать, что мы до сих пор не располагаем для этого достаточными данными. Приходится опираться лишь на весьма приблизительные расчеты (Теплинская, 1995), согласно которым в Кандалакшском заливе зоопланктонными организмами за сутки в среднем выедается 20–30% от биомассы планктонных бактерий. Конечно, распространять эти данные на все море в целом и на различные сезоны и годы преждевременно, однако они все же дают некоторое представление о том, какую роль планктонные бактерии играют как элемент питания беломорских организмов. При этом, не сомневаясь в указанных (Теплинская, 1995) величинах выедания бактериопланктона, не могу согласиться с мнением автора процитированной работы о том, что бактерии потребляются лишь зоопланктоном. Как отмечено выше, они служат источником пищи для самых разных организмов, а не только для зоопланктона.
Таким образом, можно в первом приближении подсчитать, что от общей годовой продукции бактериопланктона в Белом море, эквивалентной (см. выше) 2.4 . 1014 ккал, около 25% (т.е. 0.6 . 1014 ккал) поступает в трофические цепи в качестве питания различных консументов.
Заканчивая обсуждение роли планктонных бактерий в функционировании водных экосистем, следует отметить их участие в трансформации нефти и нефтяных углеводородов в море. По этой причине биоиндикация нефтяного загрязнения и качества воды с помощью микробиологических методов широко применяется при гидробиологическом и санитарно-техническом анализе состояния различных водоемов (Цыбань и др., 1981).
В Белом море наибольшая плотность нефтеокисляющих бактерий была зарегистрирована в вершине Кандалакшского залива, где она составляет в среднем несколько десятков (реже–сотен) клеток в 1мл воды с максимумом в верхнем 0.5-метровом слое. Плотность нефтеокисляющих бактерий быстро уменьшается по мере удаления от г. Кандалакша в открытое море, где содержание в воде нефтеокисляющих бактерий, как правило, не бывает больше нескольких кл./мл (Теплинская, 1995). В Онежском заливе они вообще не были обнаружены. Относительно высокая численность этих микроорганизмов и их заметная доля в бактериальном сообществе районов, обогащенных нефтеуглеводородами, свидетельствуют о достаточно активном самоочищении вод Белого моря от нефтяного загрязнения.



