Продукционный потенциал Белого моря
Отдельные элементы баланса вещества и энергии в Белом море, а также трофические взаимоотношения между его обитателями последовательно рассмотрены в предыдущих главах по мере того, как производилась оценка потребления пищи теми или иными организмами. Однако, это был, если можно так выразится, мелкомасштабный анализ. В этой главе будут проанализированы взаимоотношения между более крупными составляющими биоты пелагиали и бентали Белого моря. Для этого использованы результаты расчетов продукции и потребления пищи, которые для удобства дальнейшего обсуждения сведены воедино в две таблицы (табл. 35 и 36)*.
Фитопланктоном и криофлорой за счет энергии солнца, превращенной с помощью фотосинтеза в энергию химических связей синтезированных органических веществ продуцируется за год около 850 ккал в столбе воды под 1 м2. Основным потребителем их продукции являются различные организмы зоопланктона и зообентоса, отфильтровывающие микроводоросли. Бактерии, относящиеся к редуцентам, перерабатывают органические вещества и за счет энергии их химических связей строят тела своих клеток, создавая источник пищи для различных фильтраторов и детритофагов. В результате деятельности планктонных бактерий перерабатывается количество ОВ, эквивалентное энергетически 2670 ккал/м2. ____________________ * Подчеркнем еще раз, что эти расчеты весьма приблизительны, хотя бы потому, что из-за нехватки данных не учтена продукция и потребление энергии автотрофных бактерий, микробентоса, мейобентоса и макрозоопланктона (медуз и гребневиков).
Таблица 35 Продукция вещества и энергии в Белом море (по: Бергер и др., 1995 а, б, 2007; Алимов и др., 2005; Бергер, 2005, 2006 а, б, 2007; Бергер, Примаков, 2007; Примаков, Бергер, 2007)
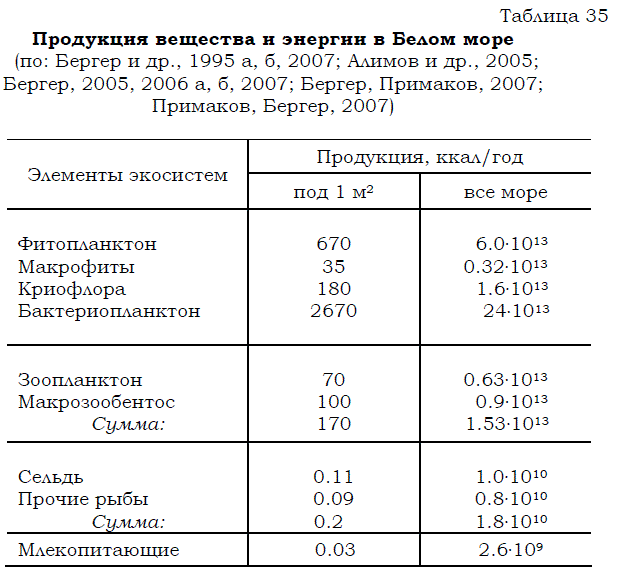
Если бы консументы первого порядка (перечисленные выше организмы зоопланктона и зообентоса) питались только за счет продукции фитопланктона и криофлоры, равной суммарно 850 ккал в столбе воды под 1м2 в год, то для них в Белом море просто не хватило бы пищи. Как следует из представленных данных (табл. 36) годовой рацион зоопланктона и макрозообентоса равен 830 ккал в столбе воды под 1м2.
Таким образом, получается, что эти консументы должны были бы выедать полностью все планктонные микроводоросли, что абсолютно невозможно. Следовательно, нехватка пищи должна быть компенсирована из другого источника. Отсюда становится очевидной та огромная роль, которую играют в трофике водоема бактерии, служащие не только деструкторами органических веществ, но и важнейшим источником пищи для многочисленных консументов, составляющих зоопланктон и зообентос Белого моря.
Таблица 36 Потребление вещества и энергии различными животными (по: Бергер и др., 1995 а, б, 2007; Алимов и др., 2005; Бергер, 2005, 2006 а, б, 2007; Бергер, Примаков, 2007; Примаков, Бергер, 2007)
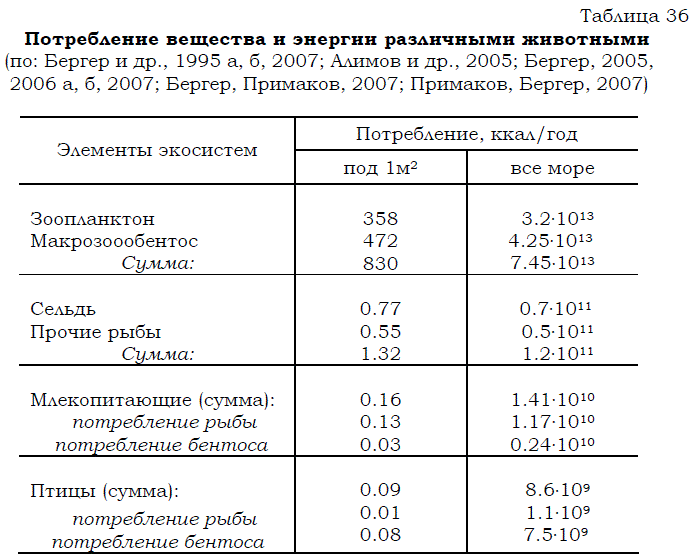
Существенный вклад в эту же копилку энергии вносят дополнительно и растворенные органические вещества, абсорбируемые через покровы тела различными моллюсками, полихетами, иглокожими и др. Оценить размеры этой составляющей по имеющимся данным пока можно лишь очень приблизительно. Было подсчитано (Andrews. Williams, 1971), что гетеротрофная утилизация растворенного органического вещества обеспечивает морским организмам количество энергии, эквивалентное половине продукции фитопланктона. Если допустить, что эти величины приемлемы и в нашем случае, тогда окажется, что за счет абсорбции РОВ беломорские бентосные и планктонные организмы получают около 340 ккал/м2. Таким образом, даже без детритной составляющей для питания организмов, входящих в состав зоопланктона и зообентоса, может использоваться количество пищи, эквивалентное примерно 4 тыс. ккал /м2. Из него за год данными консументами выедается приблизительно пятая часть: 830 ккал/м2.
Конечно, воображаемый дефицит энергии могли бы частично заполнить также и источники детрита, что действительно имеет место, однако этот процесс поступления вещества и энергии к их потребителям отличается рядом особенностей.
Во-первых, детрит в значительной степени состоит из отмирающих клеток фитопланктона, что приводит к изъятию существенной доли его продукции из пастбищной трофической цепи и не уменьшает, а только увеличивает обсуждаемый дефицит. Этот поток энергии направляется по другому каналу: она утилизируется потребителями фитопланктона не непосредственно, а через элементы детритной цепи.
Во-вторых, частицы детрита покрыты пленкой перерабатывающих их бактерий и именно в таком виде потребляются различными планктонными и бентосными детритофагами. Следовательно, представить детрит отдельно от бактерий невозможно.
Существует еще один источник детрита: макрофиты. Эти продуценты весьма обильны в Белом море и дают существенную продукцию. Ее отношение к продукции фитопланктона значительно выше такового во многих других морях Мирового океана (см. гл. 4), однако напрямую продукция макрофитов практически не потребляется, поскольку в Белом море нет организмов, питающихся макрофитами. Частично слоевища фукоидов, ламинарий и других макрофитов скоблят некоторые моллюски (например, литторины), а куски разрушающихся слоевищ поедают морские ежи. Тем не менее большая часть продукции макрофитов, в разрушении и переработке которых принимают участие бактерии, идет на образование детрита, но даже при этом она все равно не могла бы в достаточной степени компенсировать нехватку пищи для тех бентосных и планктонных организмов, о которых говорилось выше. Ее доля в продукции автотрофного звена трофической цепи ничтожно мала: 30 ккал под 1м2 за год по сравнению с 850 ккал/м2, продуцируемыми за год фитопланктоном и криофлорой (табл. 35).
Существенную роль в образовании детрита, кроме фитопланктона и макрофитов, играют и другие отмирающие организмы, которые в зависимости от размера становятся пищей детритофагов и/или трупоедов. К этому добавляются продукты жизнедеятельности различных организмов (фекалии, псевдофекалии, пеллеты и др.) и прочая взвешенная органика. Все вместе эти компоненты образуют «морской снег», оседающий на дно. Еще в толще воды в процессе оседания и по его завершении уже на дне моря эта разнородная органика частично утилизируется многочисленными сестонофагами и детритофагами. Неутилизированная часть подвергается дальнейшей переработке микроорганизмами. После минерализации эта составляющая пополняет запасы биогенных элементов, используемых впоследствии в процессах первичного продуцирования автотрофными организмами.
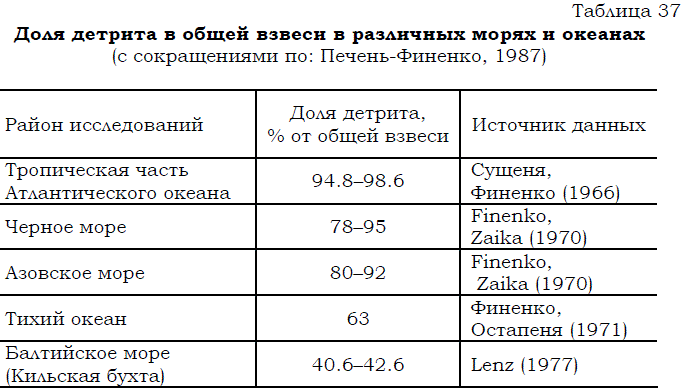
Детрит составляет самую большую после растворенной органики часть органических ресурсов моря. В нем аккумулированы значительные запасы энергии, сравнительно медленно и различными путями вовлекаемой в биотический круговорот (Печень–Финенко, 1987). Согласно схеме потоков вещества и энергии (Одум, 1986) детритная пищевая цепь служит важнейшим каналом трансформации вещества и энергии в морских и пресных водоемах. Исследования, выполненные в различных морях и океанах, показали, что на долю детрита приходится от 40 до 99% от общей органической взвеси (табл. 37).
Таблица 37 Доля детрита в общей взвеси в различных морях и океанах (с сокращениями по: Печень-Финенко, 1987)
Химический состав детрита может претерпевать значительные изменения в зависимости от сезона, характера водоема, его биоты и ряда других обстоятельств, однако во всех случаях детриту свойственно высокое содержание белков и углеводов, а также аминокислот (Мельников, 1974). Размеры детритных частиц варьируют в значительных пределах. В морских водоемах разного трофического уровня на долю фракции с размерами частиц от 10 до 200 мкм, отфильтровываемой организмами мезопланктона, приходится 15–50% от общего органического углерода взвеси. Таким образом, около половины общего количества детрита может быть использовано зоопланктонными фильтраторами (Мельников, 1974). Биомасса бактерий, ассоциированных с частицами детрита, составляет от 5 до 50% от массы самих частиц, которые бактерии обогащают белками, повышая их пищевую ценность. Установлена способность употреблять детрит (преимущественно недавно образованный) в пищу, наряду с микроводорослями, присущая как обитателям океанической пелагиали, так и неретическим видам зоопланктона. Эта способность наиболее развита у эстуарных видов планктонных ракообразных, хотя и они питаются не чистым детритом, а его смесью с планктонными микроводорослями. Таким образом, комбинация детрита и фитопланктона представляет собой единый пул энергии, более постоянный, чем каждый источник в отдельности (Печень–Финенко, 1987).
К сожалению, применительно к Белому морю пока не хватает данных для того, чтобы оценить общую продукцию планктонных и бентосных детритофагов и рассчитать, как она соотносится с продукцией отдельных звеньев пастбищной трофической цепи. Частично этот пробел можно заполнить материалами по макробентосу (А.Д. Наумов, неопубл. данные). На основании обработки этих оригинальных данных мною были построены графики изменения с глубиной суммарной продукции, а также продукции компонентов пастбищной и детритной трофических цепей в отдельности, выраженной в процентах от ее суммарной величины (рис. 28).
Представленные материалы свидетельствуют, вопервых, о том, что общая продукция макробентоса резко падает по мере продвижения от поверхности в глубину. Во вторых, наглядно демонстрируется, что аналогичная тенденция характерна и для изменения продукции компонентов пастбищной цепи. В неглубоких слоях воды (от 0 до 20 м) она составляла 99.8% от суммарной продукции организмов макробентоса, а на глубинах свыше 50 м не превышала 10%. Что касается детритофагов, то их продукция наоборот увеличивается с глубиной. Она достигает максимального уровня на глубине свыше 50 м, приблизительно в 10 раз превосходя продукцию компонентов пастбищной трофической цепи.
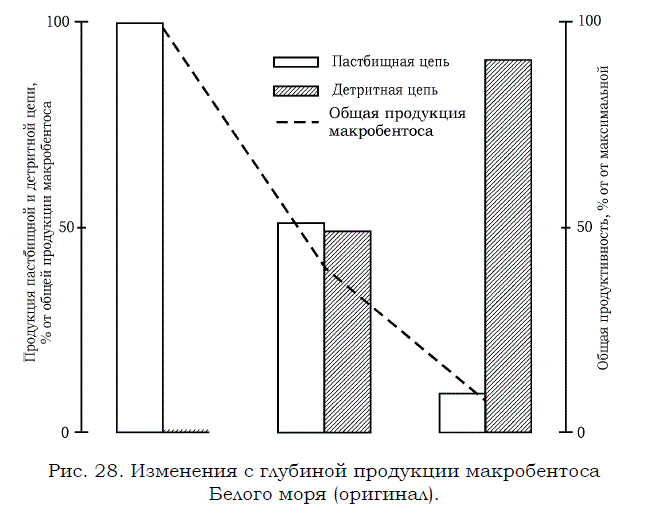
Рис. 28. Изменения с глубиной продукции макробентоса Белого моря (оригинал).
Важным элементом в балансе вещества и энергии в экосистемах водоема служат органические вещества как автохтонного, так и аллохтонного происхождения. Различные продукты жизнедеятельности беломорских организмов образуют пул относительно легко усваиваемых органических веществ. Из него черпают энергию многие организмы, абсорбирующие аминокислоты и сахара поверхностью тела, а также бактерии, редуцирующие органические вещества. Аллохтонная компонента количественно гораздо менее значима. К тому же эти вещества значительно хуже усваиваются, чем автохтонные. Если привести обе составляющие к общему знаменателю (усвояемости автохтонной органики), то окажется, что в Белое море реками за год приносится такое количество аллохтонных органических веществ, которое соответствует по усваимости 1.8 млн. т, что эквивалентно 0.9.1013 ккал (около 100 ккал в пересчете на 1 м2). Однако большая часть этого энергетического потока, эквивалентная 0.8.1013 ккал в год (около 90 ккал под 1м2), не выходит из прибрежной зоны за пределы многочисленных эстуариев, задерживаясь маргинальными фильтрами (см гл. 2). Эти органические вещества играют важную роль в эстуарных экосистемах, но оказывается, что часть из них после ряда трансформаций включается в обмен вещества и энергии в море уже за пределами эстуариев.
Эта крайне важная составляющая трофических взаимоотношений в беломорских экосистемах, заключающаяся в продуцировании и переносе вещества и энергии из прибрежных участков в более глубокие части моря, до недавнего времени оставалась вне сферы интересов исследователей. Лишь последние 20 лет ситуация начала изменяться к лучшему. Показано, что прибрежная зона характеризуется высокой продуктивностью и во многом определяет продуктивность всей акватории не только Белого, но и многих других морей (Бигон и др., 1989; Lebedev et al., 1989; Бек, 1990 б; Бурковский, 1992; Бек и др., 2000; McLusky, 2001; Barrerа-Oro, 2002 и др.).
Детальные исследования, выполненные в прибрежной части Кандалакшского залива (в эстуарии р. Черной и в губе Ярмолинской), показали (Пономарев, 2004), что сообщества рыб в этих районах представлены следующими видами:
а) в эстуарии – 2 видами колюшек Pungitius pungitius и Gasterosteus aculeatus, полярной камбалой Liopsetta glacialis, ледовитоморской рогаткой Triglopsis quadricornis и навагой Eleginus navaga; б) в губе Ермолинской к ним добавляются европейский керчак Myoxocephalus scorpius, треска Gadus morhua и европейская бельдюга Zoarces viviparus. Исходя из особенностей питания этих видов, обобщенная схема потоков органического вещества представляется следующим образом (рис. 29). Благодаря материковому стоку, прибрежные воды обогащаются органическим веществом. Интенсификация стока в весенне-летний период способствует массовому развитию фито-, а затем и зоопланктона, служащего кормом для молоди практически всех рыб Белого моря. В это время при обильном стоке органики и развитии планктона наблюдается массовый заход трехи девятииглой колюшки в устья рек, где происходит их нерест.
Будучи высокоэвригалинными видами, эти рыбы могут проникать вглубь эстуариев, где становятся не доступными для менее эвригалинных хищников (рогатка, керчак, навага и треска). Их потомство также растет и развивается за счет ресурсов этой части прибрежной зоны, не доступной для других видов. К концу лета взрослые колюшки и их молодь откочевывают из максимально опресненных прибрежных зон в более мористые и глубокие районы (Мухомедьяров, 1966), где становятся доступными для других видов рыб. Таким образом, оба вида колюшек являются своего рода «накопителями» и «переносчиками» органического вещества и энергии из наиболее опресненных частей эстуариев, не доступных для других видов рыб, в открытые районы моря, с последующим включением этого вещества в трофические цепи (рис. 29,А). Рост численности колюшки, как уже отмечалось ранее, наблюдающийся в последнее время (Иванова и др., 2007), приводит к интенсификации процессов транспорта энергии и вещества из опресненных прибрежных районов вглубь Белого моря.
В менее опресненной части прибрежной зоны (губа Ермолинская, мористые участки эстуария р. Черной) поступление органического вещества со сточными водами ручьев при таянии снега и за счет деструкции макрофитов также во многом обеспечивают существование сообществ обитателей этих участков акватории (рис. 29, Б). Структура ихтиоценозов в этой зоне усложняется: повышается видовое разнообразие ихтиофауны. Если в летний период здесь откармливаются полярная камбала, треска, навага и керчак, то к зиме большинство из них откочевывает на глубину, а в прибрежной зоне зимуют только рогатка и бельдюга (Андрияшев, 1954). Таким образом, многие беломорские рыбы, откармливающиеся летом в прибрежной зоне, во время зимних миграций на глубину обеспечивают транспорт в пелагиаль органического вещества, продуцированного в прибрежье. Рыбы тех видов, которые остаются зимовать в этой зоне, оказываются вовлеченными в локальные циклы трансформации органического вещества.
Значимость процессов обмена веществом и энергией между прибрежными, в том числе и эстуарными экосистемами, с одной стороны, и экосистемами открытого моря, с другой, очевидна, и ее трудно переоценить. Благодаря им, не только автохтонные ОВ, произведенные в прибрежных участках моря, но и аллохтонная органика, приносимая реками и большей частью остающаяся в прибрежной зоне из-за деятельности «маргинальных фильтров», вовлекаются в круговорот вещества и энергии всего моря. Эти интеграционные процессы, несомненно, способствуют максимальной реализации продукционных возможностей всех беломорских экосистем. К сожалению, несмотря на результаты исследований, описанных выше, приходится в очередной раз отметить слабую изученность рассматриваемых процессов, как и многих других составляющих биоэнергетического баланса Белого моря.
Описанный феномен не есть нечто уникальное, присущее только Белому морю. Наоборот, показано (BarrerаOro, 2002), что в приантарктических водах прибрежные рыбы играют ту же роль накопителя и переносчика вещества и энергии от прибрежья в глубокие участки моря. В связи с этим можно, по-видимому, предполагать, что такая организация взаимоотношений между экосистемами прибрежья и открытого моря достаточно универсальна, однако было бы преждевременно утверждать, что она распространяется на все районы Мирового океана: для этого пока не хватает данных.
Благодаря утилизированным веществу и энергии, планктонные и бентосные животные, кроме покрытия трат на обмен, производят продукцию, величина которой за год во всем море равна (табл. 35) соответственно 0.63.1013 (70 ккал под 1 м2) и 0.9.1013 (100 ккал/м2), а в сумме составляет 1.53.1013 ккал (170 ккал/м2). Эта продукция в виде объектов питания в свою очередь утилизируется консументами более высокого порядка, и в первую очередь рыбами. Последних можно разделить на две основные группы по характеру питания и той роли, которую они играют в функционировании беломорских экосистем и промысле: планктонофагов и бентофагов. Количество рыб, имеющих преимущественно хищный тип питания, относительно невелико (Расс, 1995).
Основу ихтиофауны и промысла, как уже неоднократно отмечалось, составляют зоопланктонофаги. К ним относятся сельдь, мойва, навага, корюшка и трехиглая колюшка. При этом следует заметить, что среди перечисленных рыб во взрослом состоянии преимущественно зоопланктоном питаются лишь сельдь, мойва и трехиглая колюшка. Остальные потребляют организмы зоопланктона лишь в течение относительно короткого периода: при переходе на активное питание и некоторое время после этого, а во взрослом состоянии питаются в основном бентосными организмами (Подражанская, 1995). Пищевой конкуренции между сельдью и остальными названными выше видами практически нет. Среди трех основных рыбзоопланктонофагов наиболее значима сельдь, на долю которой приходится наибольшая биомасса (в настоящее время – около 12 тыс. т).
Если соотнести годовую продукцию зоопланктона (0.63.1013 ккал во всем море или около 70 ккал в столбе воды под 1 м2) с его потреблением сельдью (0.7.1011 ккал в расчете на все море или 0.77 ккал под 1 м2), то оказывается, что сравниваемые величины различаются практически на два порядка. Отсюда логично сделать вывод о том, что запасы зоопланктона используются лишь в малой степени (около 1% от продукции). Следовательно, запасы сельди в Белом море не лимитированы пищевыми ресурсами экосистем пелагиали и могут быть значительно больше нынешних, которые равны приблизительно 12 тыс. т.
Однако этот вывод не дает ответа на вопрос о том, сколько же сельди может прокормить море? Рассчитать возможные запасы сельди (и других зоопланктонофагов, в том числе таких рыб, как мойва и трехиглая колюшка) можно, исходя из уровня продукции фитопланктона. Калорийность морского фитопланктона составляет 0.25 ккал/г (см. табл.1). Следовательно, при годовой продукции фитопланктона 6. 0 13 ккал приращение его биомассы составляет порядка 2.4.10 14 г. При соблюдении правила 10%-ной экологической эффективности (Elton, 1947; Раймонт, 1983) годовое приращение биомассы консументов I порядка (зоопланктона) должно соответствовать 2.4.1013 г, а увеличение биомассы зоопланктонофагов – 2.4.1012 г.
Это означает, что если бы сельдь была единственным зоопланктонофагом в Белом море, то запасы зоопланктона, исходя из результатов этого расчета, могли бы обеспечить продукцию сельди в объеме приблизительно 2.5 млн. т в год. При годовом Р/В-коэффициенте рыб, равном 0.45, такая продукция соответствует запасам сельди около 5 млн. т.
Очевидно, однако, что такое допущение некорректно. Зоопланктоном, кроме сельди, питаются другие рыбы, а также пелагические и бентосные кишечнополостные: гребневики, медузы, различные гидроидные и коралловые полипы (горгонариии и актинии) и некоторые другие беспозвоночные. Среди всех этих зоопланктонофагов после рыб основными потребителями зоопланктона можно в первую очередь назвать сцифоидных медуз и гребневиков. Их численность в Белом море в летние месяцы может быть весьма значительной, но, к сожалению, данных об обилии этих беспозвоночных до сих пор крайне мало, прежде всего изза отсутствия надежных количественных методов их учета. Что касается питания медуз и гребневиков, то до сих пор исследован в основном состав их пищи, а ее количество гораздо менее изучено, во всяком случае применительно к обитателям Белого моря.
Среди имеющихся работ этого направления исследований следует в первую очередь назвать публикации В.А. Свешникова (1963) о питании беломорских медуз, Н.П.Логиновой и Н.М. Перцовой (1967) о питании гребневиков и сцифомедуз и кандидатскую диссертацию А.А. Прудковского (2006), посвященную изучению питания массовых гидромедуз Белого моря. Общий вывод, вытекающий из результатов всех этих работ, заключается в том, что массовые виды кишечнополостных в Белом море могут составлять существенную пищевую конкуренцию сельди, поедая значительное количество зоопланктона.
Убедительно показано, сколь велико количество пищи, потребляемое отдельными особями. Так, например, одна сцифомедуза Aurelia aurita в зависимости от своих размеров съедает за сутки от 40 до 187 мг зоопланктона (Логинова, Перцова, 1967). При этом, однако, не установлено, сколько времени питаются медузы и какова их плотность, что лишает возможности оценить выедание зоопланктона этими животными.
По данным тех же исследователей один гребневик Bolinopsis infundibulum потреблял в сутки в среднем 0.9 мг пищи, а в их скоплениях все питающиеся особи выедали около 48.8 мг зоопланктона из 1 м3 воды. Конечно, если считать, что средняя бимасса зоопланктона в Белом море составляет, как было показано выше, 200 мг/ м3, то можно заключить, что одни только гребневики данного вида выедают около 25% биомассы зоопланктона. Но считать так было бы ошибкой по нескольким причинам:
- Пик численности гребневиков приходится только на июль–начало августа, а в остальное летнее время встречаются лишь единичные экземпляры. 2. Гребневики распространены в основном на мелководье. Пик их численности приходится на глубины порядка 5 м (Логинова, Перцова, 1967). 3. Подсчитанная плотность скоплений этих животных не позволяет сказать что-то более или менее определенное о средних показателях их обилия. Следовательно, реально оценить пресс этих зоопланктонофагов пока нельзя. Можно лишь повторить, что они могут быть серьезными пищевыми конкурентами сельди, по крайней мере, в течение 1-2 летних месяцев и в основном в мелководных прибрежных районах Белого моря, а сельдь, как известно (см. гл.5), в летнее время откармливается главным образом в Бассейне, т.е. далеко от берегов. Кроме того, ситуация не ограничивается только конкуренцией за пищу: и гребневики, и сцифомедузы могут в некотором количестве поедать икру и молодь рыб – сельди, мойвы и некоторых других видов (Southward, 1955; Камшилов, 1960; Свешников, 1963; Логинова, Перцова, 1967; Раймонт, 1983 и др.).
Существенную пищевую конкуренцию сельди и другим ихтиопланктонофагам могут составить и массовые беломорские гидромедузы, которые в период максимального развития выедают около 10% зоопланктона (Прудковский, 2006). Кроме того, гидроидные полипы и гидромедузы в период максимальной численности могут уничтожать большое число личинок сельди, погибающих от контакта с их стрекательными клетками, что было установлено экспериментально (Иванченко, 1995).
Конечно, эти фрагментарные сведения не позволяют оценить потребление беломорского зоопланктона всеми зоопланктонофагами и вычленить из него долю сельди. Потребление зоопланктона – предмет конкурентных взаимоотношений различных консументов, между которыми устанавливается то или иное динамическое равновесие. На данный момент оно, по-видимому, сдвинуто в сторону не сельди, а иных потребителей зоопланктона. Последнее не означает, тем не менее, что сельдь ограничена в питании. Более высокоорганизованные, активные и подвижные рыбы легко могут выдержать конкуренцию за пищу с другими зоопланктонофагами (медузами и гребневиками), быстро находя скопления зоопланктона и реагируя на его горизонтальные и вертикальные перемещения. Существенным конкурентом сельди в этом отношении в ближайшее время станет (если уже не стала) трехиглая колюшка, количество которой резко возросло в последние годы (Иванова и др., 2007). Однако, несмотря на все эти оговорки, можно попрежнему утверждать, что беломорская сельдь не испытывает ограничений в количестве пищи, и ее запасы могут быть значительно (не менее, чем на порядок) больше нынешних.
Кроме того, можно сделать еще один вывод. Суть его заключается в том, что в нынешней ситуации, когда запасы сельди крайне малы, продукция зоопланктона потребляется в основном медузами и гребневиками, для которых создались благоприятные кормовые условия. Если при этом в Белое море тем или иным способом (например, с балластными водами судов) проникнет какой-нибудь вселенец– зоопланктонофаг (например, гребневик или медуза), ситуация может стать неуправляемой и вызовет катастрофические последствия не только для пелагических экосистем, но и для моря в целом. Подобное предположение – не чисто гипотетический вариант. Такие случаи хорошо известны. Так, например, отсутствие хищников и паразитов в новом биотопе создало предпосылки для интенсивного развития популяции вселенца – гребневика Mnemiopsis leidyi, который питается рачковым планктоном, икрой и личинками рыб. Его появление (в 1980 г.) и последующее массовое развитие в Черном, Азовском и Каспийском морях в последние десятилетия привели к подрыву кормовой базы рыбпланктонофагов и резкому падению их вылова (Шиганова и др., 2003; Виноградов, 2006; Комакин, Егоров, 2006). Недавно гребневики этого вида были обнаружены впервые сначала в Балтийском (Javidpour et al., 2006), а затем и в Северном море (Boersma et al., 2007).
Подобные случаи инвазии – феномен достаточно обычный для различных организмов, не только гребневиков. Не останавливаясь подробно на обсуждении этих вопросов, отметим, что такие случаи зарегистрированы и в Белом море, где, например, недавно в эстуарии Северной Двины был обнаружен китайский мохнорукий краб Eriocheir sinensis (Бергер и др., 1999; Berger, Naumov, 2002; Наумов, Бергер, 2004). Этот вселенец проник в Белое море из Балтики, мигрируя по водоемам бассейнов обоих морей и преодолевая по суше расстояния между отдельными озерами и реками.
Если ситуация с «желейным» макрозоопланктоном пока остается лишь предполагаемой, то резкое увеличение численности трехиглой колюшки в Белом море в последние годы – реальный факт. Нынешнее обилие этого вида обусловлено, на наш взгляд, двумя причинами:
1) частичным восстановлением зарослей зостеры, в которой колюшки делают свои гнезда и выводят молодь; 2) обилием зоопланктона, являющегося основным
источником пищи для трехиглой колюшки. Рост обилия трехиглой колюшки вызывает существенные изменения не только путем повышения пищевой конкуренции среди планктонофагов. Не менее важно, что при этом происходят (или должны происходить, но пока еще не подтверждены фактическим материалом) многие другие экосистемные перестройки.
Колюшка, как уже сообщалось выше, является объектом питания многих рыб Белого моря (трески, бычков, сельди, поедающей в числе прочего и молодь колюшки, и некоторых других видов рыб). Неурожайные для колюшки годы сильно сказываются на запасах промысловых рыб (Подражанская,1995).
Следовательно, есть все основания ожидать, что увеличение обилия трехиглой колюшки, наблюдающееся самые последние годы, должно в первую очередь отразиться соответствующим образом на изменении ее доли в рационе рыб ряда видов. Такие факты начинают накапливаться.
По неопубликованным и еще подлежащим уточнению данным П.Н. Ершова летом (июль–август) 2007 г. трехиглая колюшка была обнаружена в желудках 54.8% вскрытых экземпляров крупной трески (длина тела не менее 30 см), пойманных в губах Чупа и Кив в Кандалакшском заливе. Доля колюшек в пищевом комке составляла 53% от массы съеденной пищи. При этом число трехиглых колюшек, обнаруженных в желудках одной треской, менялось от особи к особи в пределах от 1 до 34 экз.
Исследования, проведенные в этом же районе Белого моря осенью предыдущего года, показали (Фукс, 2007), что трехиглая колюшка не входила в число 19 пищевых объектов, обнаруженных в желудках трески*. Сравнение результатов этих двух работ свидетельствует о том, что пищевые отношения трески достаточно пластичны и очень быстро перестраиваются, реагируя на изменения ситуации в экосистемах моря. Подобная пластичность питания характерна и для других рыб (Подражанская, 1995).
Аналогичные изменения затрагивают и морских птиц. Трехиглая колюшка играет особенно важную роль в питании самой многочисленной из морских птиц Белого моря – полярной крачки. Как показано многолетними наблюдениями (Бианки и др., 1995), массовое гнездование крачек возможно в Кандалакшском заливе при значительных подходах трехиглой колюшки к берегам. В 60–90-х годах прошлого столетия, когда численность колюшки была очень низкой, крачки гнездились в малом числе. В наши дни ситуация с обилием полярной крачки должна измениться на противоположную. Такой прогноз представляется достаточно реалистичным, однако произойдет это или нет, и если произойдет, то насколько быстро, покажет будущее.
Вторая группа рыб (бентофаги) базируется в своем питании на продукции макрозообентоса, составляющей за год ____________________ * Размеры исследованных рыб не указаны. (Прим. автора).
во всем море около 0.9.1013 ккал (100 ккал/м2). Из этого рыбами-бентофагами в течение года потребляется всего 0.5.1011 ккал (0.55 ккал /м2), т.е. менее 0.5% от продукции зообентоса. Даже если к этому добавить годовое выедание бентосных организмов птицами (0.75.1010 ккал в расчете на все море или 0.08 ккал/м2) и морскими млекопитающими (0.24.1010 ккал или 0.03 ккал/м2), то и тогда эта суммарная величина, равная 6.0.1010 ккал (0.66 ккал/м2), будет меньше 1% от годовой продукции донных беспозвоночных.
Конечно, при этом не следует забывать, что из наших расчетов полностью исключены многие виды рыб (бычки, маслюк, бельдюга и др.): их запасы совершенно не исследованы. Однако, если даже предположить, что они не меньше таковых у ранее названных рыб-бентофагов, то и в этом случае потребление бентоса будет слишком мало, чтобы лимитировать развитие бентосоядных рыб в Белом море.
Следовательно, эта группа рыб также не лимитирована имеющимися запасами зообентоса и его продукцией.
Как уже неоднократно отмечалось (см. гл. 2) в Белом море в летнее время существенно прогреваются лишь верхние слои воды. На глубинах более 50–60 м температура воды не поднимается выше 0°С в течение всего года. В связи с этим большие глубины, занимающие центральную часть моря, на долю которых приходится почти половина площади всей акватории, оказываются не пригодными для основных донных и придонных рыб (навага, треска, камбаловые, зубатка и корюшка). Благоприятны для их откорма мелководья, зона которых наиболее обширна в заливах, но и на этих участках значительные площади имеют низкую кормность. Кроме того, большая доля биомассы бентоса в Белом море приходится на моллюсков, в первую очередь двустворчатых, а они – основа корма лишь немногих видов, в первую очередь таких специализированных зообентофагов, как зубатка и камбаловые. Многие другие бентосоядные рыбы потребляют лишь мелких моллюсков, да и то в небольших количествах. Они предпочитают питаться полихетами и ракообразными.
Таким образом, хоть биомасса беломорского бентоса и его продукция достигают значительных величин, но его кормовое значение относительно невелико (Кудерский, 1966; Кудерский, Скарлато, 1986). По-видимому, именно по этой причине многие рыбы-бентофаги в больших количествах потребляют в качестве дополнительных источников питания различных рыб, имеющих гораздо большую питательную ценность (калорийность), чем бентосные организмы (Подражанская, 1995; Тарвердиева, 2000).
Определение уровня рыбопродуктивности – одна из важнейших задач (если не самая важная), которую необходимо решить для организации рационального использования биоресурсов интересующих нас водоемов. Попыток ее решения не так много.
Анализ многочисленных данных, произведенный Г.Г. Винбергом и его учениками, соавторами и последователями (Winberg, 1972; Бульон, Винберг, 1981; Алимов, 1989, 2004; Бульон, 1994), показал, что уровень вылова рыбы в различных пресных водоемах составляет обычно около 0.1–0.12% от продукции фитопланктона. В некоторых озерах и водохранилищах он повышается до 0.3%, а максимального уровня (0.4–2.2%) достигает в прудах.
Подобный анализ в отношении морских водоемов базируется на гораздо меньшем фактическом материале. В.В. Бульон и Г.Г. Винберг (1981) рассчитали, что в Мировом океане вылов рыбы составляет наименьшую долю от продукции фитопланктона: 0.01–0.02%. По их данным в таких внутренних морях, как Балтийское, Азовское, Черное, Каспийское, Аральское и залив Мэн в Атлантическом океане, этот показатель не превышает 0.1%. П.А. Моисеев (1969) рассчитал, что в морях уровень вылова рыбы не превышает 0.04–0.05% от величины первичной продукции*. В Охотском море в годы максимального вылова рыбы это соотношение выражалось величиной около 0.02% (Шунтов, 1985). Близкие данные (0.03%) для некоторых морей привел в своей работе и В.В. Бульон (1994). Таким образом, по данным разных авторов и для разных морей рассматриваемый показатель варьирует от 0.02 до 0.1% при средней величине около 0.06%.
В соответствии с этими представлениями можно рассчитать возможный годовой вылов рыбы в Белом море. Он эквивалентен энергетически 1.2.1010–6.0.1010 ккал при сред____________________ * Имелась в виду продукция фитопланктона. (Прим. автора).
ней величине около 3.6.1010 ккал. Если исходить из средней калорийности рыб около 1.1 ккал/г, то их общий вылов в Белом море должен составлять в среднем около 32 тыс. т в год. При самых низких и самых высоких величинах коэффициента, используемого для анализа, вылов рыбы будет составлять соответственно 11 и 54 тыс. т в год, однако такой расчет приемлем лишь в том случае, когда основу вылова составляют рыбы средней калорийности. Что касается беломорских промыслов, то добыча сельди в периоды их максимального расцвета (табл. 38) составляла около 80% от общего вылова рыбы. Если принять это соотношение разных видов рыб в промысле за основу, то получится, что уловы сельди, рассчитанные, исходя из ее более высокой калорийности и среднего значения используемого коэффициента, могли ежегодно составлять около 15 тыс. т. При минимальном и максимальном значении этого коэффициента они должны быть равны соответственно 5 и 26 тыс. т.
Таким образом, на основании результатов этих расчетов можно сделать вывод о том, что вылов рыбы в Белом море может быть больше нынешнего примерно в 10 раз.*
К этому следует добавить еще один вывод. Повидимому, максимальные уловы рыбы, достигавшие примерно 170 лет назад суммарно около 40 тыс. т в год, из которых на долю только сельди приходилось 32–34 тыс. т (табл. 38), превышали допустимый уровень изъятия. В основном это касалось беломорской сельди. Можно предполагать, что столь значительный «перелов» как раз и привел к первоначальному подрыву ее запаса, в то время, по-видимому, максимального.
________________ * Все построения базируются на одном фундаментальном предположении, по сути дела никем и никогда не подтвержденном. Оно заключается в априорной уверенности, что продуктивность основных элементов экосистем пелагиали и бентали моря в период максимальных промыслов (т.е. примерно полтора – два столетия назад) была такой же, как сейчас. Основанием для такого предположения могут (но лишь косвенно) служить обсуждавшиеся ранее (см. гл. 2) результаты многолетнего (в одном случае 200-летнего) климатического мониторинга и данные 50-летнего круглогодичного мониторинга видового разнообразия и показателей обилия зоопланктона, а также основных гидрологических параметров в устьевой части губы Чупа.
В свою очередь это послужило одной из первоначальных причин резкого снижения уровня промысла сельди (и вообще всей рыбы) в Белом море.
Таблица 38 Максимальный уровень промысла в Белом море (по: Житний, 2007)

*В таблице не указаны наблюдавшиеся эпизодически максимальные уловы сайки (8 тыс. т в год) и пинагора (1.5 тыс. т в год), о которых говорилось ранее (см. гл.5).
Помимо бентосных организмов, выедаемых за год птицами (0.75.1010 ккал в расчете на все море или 0.08 ккал под 1 м2) и морскими млекопитающими (0.24.1010 ккал – на все море или 0.03 ккал/м2), эти животные, находящиеся на вершине пищевой пирамиды, выедают большое количество рыбы. Популяциями обитающих в Белом море морских млекопитающих (кольчатой нерпы и белухи) за год выедается количество рыбы, эквивалентное 1.17.1010 ккал (см. табл. 36). Общий запас рыб соответствует энергетически 4.0.1010 ккал, а их годовая продукция эквивалентна 1.8.1010 ккал (см. табл. 35). Следовательно, морскими млекопитающими за год выедается количество рыбы, составляющее 65% от продукции или 29% от запаса рыб.
На долю птиц-ихтиофагов приходится в год около 1 тыс. т съеденной рыбы (см. табл. 36), что в энергетическом выражении в расчете на все море равно 1.1.109 ккал и составляет около 3% от запаса и 6% от годовой продукции рыб. Для сравнения укажем (табл. 39), что в последние годы учтенный вылов рыбы в Белом море, не превышающий 2 тыс. т в год, составляет около 6% от запаса рыб и 12% от ихтиопродукции. 222
Таблица 39 Потребление рыбы в Белом море
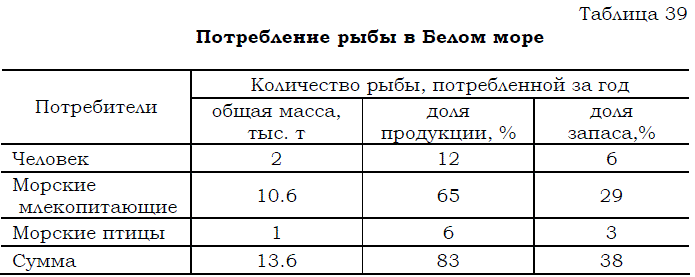
Анализируя приведенные материалы (табл. 39), не следует забывать, что доли продукции и запасов рыб, изымаемых людьми из Белого моря, рассчитаны на основании только учтенных уловов, сведения о которых соответствуют данным промысловой статистики (Отчет СевПИНРО, 2003; Доклад..., 2004, 2005; Житний, 2007). Как уже неоднократно подчеркивалось, эти данные не учитывают объем местного (любительского) вылова рыбы, который по различным экспертным оценкам (Яковенко, 1995 и др.) может быть в 2–3 раза выше указанных выше величин, однако и в этом случае, при увеличении вылова в 3–4 раза, доля вылова рыбы человеком будет меньше суммарного потребления рыбы морскими птицами и млекопитающими.
На основании результатов проведенного сравнения сам собой напрашивается лежащий на поверхности вывод о том, что хорошо бы уменьшить долю морских млекопитающих и птиц в добыче рыбы, сведя их количество к минимуму, и тогда уловы значительно возрастут. К сожалению, подобных примеров «борьбы с природой» и ее переделки на памяти человечества значительно больше, чем хотелось бы. (Вспомним хотя бы историю уничтожения бизонов в Америке и воробьев в Китае. Ни к чему хорошему это не привело, да и не могло привести). Экологическое равновесие нельзя сдвинуть и нарушить искусственными воздействиями на экосистемы без катастрофических последствий. Тем не менее доля человека в вылове рыбы в Белом море может быть постепенно увеличена. При этом подобное увеличение должно естественным образом следовать (но не опережать) за повышением запасов рыбы в результате принятых мер по устранению нарушений их воспроизводства и организации рационального промысла.
Невольно возникает еще один вопрос: почему на Белом море не вылавливается даже то количество рыбы, которое могло бы ежегодно добываться, исходя из нынешнего состояния запасов и современных способов регулирования промысла путем определения общего допустимого улова – ОДУ (табл. 40)?
Таблица 40 Общий допустимый улов (ОДУ) и уровень промысла (с изменениями по: Отчет СевПИНРО, 2003; Житний, 2007)
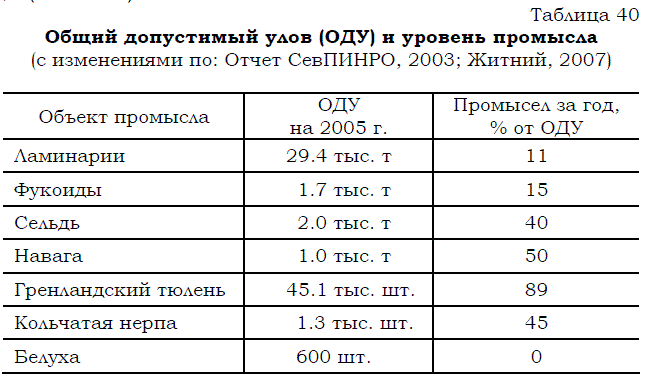
Тот же самый вопрос можно задать не только в отношении вылова рыб, но и применительно к промыслу водорослей и морских млекопитающих в Белом море.
Ответ на эти вопросы можно найти, проанализировав состояние промысловой базы и общую демографическую ситуацию на Белом море. Сделать это помогают, в частности, материалы ФГУ «Карелрыбвод» о том, как в течение ХХ– начале XXI вв. изменялся промысел рыбы по Карельскому берегу от г. Кемь до границы Мурманской области (Липатов, 2007). Показано, что с 1900 по 1910 гг. от Кеми до Ругозерской губы располагалось 160 сельдяных, сиговых и сёмужьих тоней. С 1927 г. и до середины 80-х годов прошлого столетия база промысла росла и совершенствовалась. Внедрялись тралы, кошельковые, закидные и ставные невода, кильноты, лампары, дрифтерные сети, мерёжи и др., что привело к укрупнению участков. Традиционные орудия лова (тайник, убег, гарва и др.) тоже применялись. Промысел вёлся по всему побережью и в открытом море (218 участков в 1952 г.). За последние 15 лет промбаза резко сократилась. Применяются сети, наважьи и сельдяные заколы и мерёжи. Лов ведётся на 46 участках (всего около 70 тоней). Промысел морских млекопитающих полностью прекратился с 1996 г., хотя лимиты на их добычу выделяются.
Таким образом, в указанном районе беломорского побережья в течение столетия наблюдалось изменение распределения промысловой базы от равномерного с традиционными орудиями лова к сосредоточенному на наиболее уловистых участках с применением высокоэффективных орудий. В настоящее время промысел угнетён. Б?льшая часть побережья не используется, качественный состав уловов изменился в сторону снижения доли лососёво-сиговых видов рыб.
Не останавливаясь подробно на дальнейшем анализе этой проблемы, отметим, что она детально рассмотрена рядом исследователей (Стасенков, 1995; Зеленков и др., 1995; Отчет СевПИНРО, 2003; Житний, 2007 и др.), указывающих в качестве одной из основных причин нехватку рыбаков и снижение промысловых усилий. Согласно имеющимся данным (Якобсон, 1914) количество рыбаков на участке от дер. Поньгома до устья р. Поной в период перед Первой Мировой войной превышало 9 тыс. человек. В целом на всем побережье Белого моря их было около 30 тысяч (Житний, 2007). В наше время число беломорских рыбаков, участвующих в промысле (без учета любительского лова рыбы), не превышает 2 тыс. человек. Резко сократилось и число орудий лова, используемых на промысле сельди, наваги и других рыб (Стасенков, 1995; Зеленков и др., 1995; Отчет СевПИНРО, 2003; Житний, 2007 и др.).
В целом, проанализировав динамику количества рыбаков на Белом море и изменения числа выставляющихся орудий лова, указанные выше авторы пришли к следующим выводам: 1. Число рыбаков, участвующих в промысле сельди, наваги и других рыб, примерно за 100 лет сократилось в десятки раз. 2. Сходным образом изменилось число используемых орудий
лова. 3. Многие промысловые организации (например, рыболовецкие колхозы «Север», «Северный полюс» и другие) из года в год сокращали объем добычи тех или иных объектов или вовсе прекратили его в последнее время по экономическим причинам.
Отмеченные тенденции изменения беломорских промыслов совпадают с демографической и социальноэкономической ситуациями в регионе. Всего за 20 лет (с 1950 по 1970 гг.) численность населения поморских деревень сократилась в 3 с лишним раза. В последующие 12 лет она уменьшилась еще на 28% по отношению к численности населения в 1970 г. (Васильев, Куранов, 1985). Многие рыболовецкие фактории, промысловые участки и тони заброшены. Большинство поморских деревень (Кереть, Семжа, Соностров и многие другие), существовавшие сотни лет, уже прекратили свое существование или медленно вымирают. В тех деревнях, которые еще как-то держатся, все больше пожилых людей и все меньше детей и молодежи. Из-за отсутствия работы и плохих жилищных и бытовых условий молодые люди перебираются в города, покидая берега Белого моря навсегда или возвращаясь на родину лишь на короткий период во время летних отпусков.
Исправить эту ситуацию можно только с помощью комплексной системы мероприятий, направленных на восстановление запасов и промысла основных объектов, изменения условия работы и быта рыбаков и их семей, повышения закупочных цен на добываемую рыбу, морского зверя и водорослей, развития иных видов местных промыслов и ряда других социальноэкономических мер. Все это вместе взятое требует прежде всего коренного изменения отношения не только государства, но и каждого из нас к некогда богатому и процветающему краю, каким было Беломорье всего лишь 100–150 лет назад.
В качестве одной из основных мер, направленных на восстановление уровня беломорских промыслов и создание новых рабочих мест для населения, может быть названо развитие марикультуры различных рыб (сельди, радужной форели, горбуши и, возможно, зубатки и кумжи) и водорослей (прежде всего – ламинарии сахаристой). По-видимому, к этому списку можно будет в дальнейшем добавить и другие объекты, если будут отработаны методы их культивирования, однако пока биотехнология марикультуры разработана (и в той или иной степени апробирована на практике) лишь для названных выше видов.
Все эти вопросы будут вкратце обсуждены в «Заключении» при анализе путей выхода из промысловой депрессии и восстановления промыслового потенциала Белого моря.



