Общая характеристика литорали
Литораль — граница трех сфер: лито-, гидро- и атмосферы, Будучи частью морского дна, она непосредственно примыкает к суше и находится под влиянием материкового стока, опресненного и насыщенного органикой. Как любой другой биотоп, она трехмерна, т. е. имеет длину (вдоль линии прилива), ширину (перпендикулярно первой вплоскости дна) и толщину, складывающуюся из обитаемого слоя осадка и слоя воды над дном. Более чем любой другой биотопона подвержена временным изменениям (приливо-отливным, сезонным). Особенности всех этих характеристик на отдельных участках зависят от их геоморфологии, являясь результатом длительного взаимодействия суши, моря, климата.
Разнообразие биономических типов литорали может быть сведено к принятой в отечественной литературе классификации (Кусакин, 1977). Широко распространенный умеренно защищенный берег (биономический тип I), открытый океанический берег (тип III) Воронки и Горла, эстуарии (тип V) и ванны (тип VI) характеризуются всем набором основных признаков. Биономическому типу II (открытый морской берег со значительной прибойностью) отвечают обширные пространства практически не обитаемой осушной полосы Терского берега, Двинского и части Онежского заливов. Однако такие характеристики типа II, как повышенная, приближенная к океанической, соленость и значительная аэрация, для этих побережий необязательны, в чем отражается специфика моря сего пониженной соленостью и интенсивным течением окислительных процессов (Черновская, 1956). Полностью или почти полностью выпадает биономический тип IV (лагуны).
Обитаемую литораль умеренно защищенных берегов, характерную, например, для Кандалакшского залива, можно в самом общем виде классифицировать следующим образом: мысы и другие подверженные волнобою участки чаще всего имеют скалистую и каменистую литораль, небольшую по ширине, наклонную, губы и затишные участки — илистую, илисто-песчаную и песчаную литораль, широкую и пологую. Ббльшая часть приливо-отливной зоны представляют собой переходные валунно-песчаный, валуннопесчано-глинистый и валунно-песчано-илистый типы с разным углом наклона и разной ширины. Рассматриваемая литораль, как правило, чрезвычайно пестра в силу расчлененности берега и сочетания на небольших пространствах разных фаций,
Систематические колебания приливо-отливного уровня определяют вертикальную зональность литорали. Она обусловлена как продолжительностью осушения, так и рядом факторов, опосредованных через воду: соленостью, освещенностью, гидростатическим давлением, гранулометрической дифференцировкой осадка, высеиванием органических частиц, в том числе коагулирующего растворенного органического вещества (РОВ), положением зоны LLLLaCCCCCCS LLCCCLLCLLLCLLLLCCT 0LLLCLLLLC LaLaLLLLC LLLLLLLLS нием мертвой зоны — зоны смерзания подошвы ледового припая вверхней части литорали с грунтом, а также и действием биологических факторов. Их градиент имеет тот же вектор, что и приливо?TIIHBHEle Te?e HH H. - перпендикулярный K береговой THHH HI HJIH широтный — в системе обозначений (длина, ширина и толщина литорали), принятойнами выше. Это в ряде случаев позволяет использовать для описания схему Вайана, поделившего литораль на горизонты и этажи по границам сизигийных, квадратурных и средних уровней приливов и отливов. Вертикальная зональность биоты не всегда совпадает с зональностью среды по схеме Вайана, так как она обусловлена разнообразием локального сочетания вышеперечисленных факторов (интегральным широтным векTopo M).
Несмотря на амфибиотичность условий, складывающихся на литорали, она заселена преимущественно морскими организмами. Лишь у верхней ее границы появляются обитатели суши — OEPCII (3 BOH OHH ble | IOL CTIH. KH. Растительность приливо-отливной зоны представлена в основном фукоидами. Напологой литорали в теплое время года могут долго вегетировать нитчатки. Высшие растения — галофиты — типичны на песке в верхней трети литорали, в нижней литорали на песке и иле развивается зостера. В микрофитобентосе преобладают диатомеи (Возжинская, Блинова, 1971; Бондарчук, 1974). Известное к настоящему времени животное население литорали насчитывает приблизительно 120 видов макро-, 200 видов мейо- и 500 видов микробентоса.
Среди видов, населяющих приливо-отливную зону, лишь небольшая часть принадлежит только ей, поэтому целесообразна экологическая классификация, выделяющая три группы.
1. Собственно литоральная фауна, весь жизненный цикл которой развертывается на осушной полосе (например, виды родов Gamma rus H Marinogamma rus, Jaera albifrons H Jnp. ).
2. Фауна, общая для литоралии сублиторали или литоралии супралиторали, которую в свою очередь можно разделить на две подгруппы: а) виды, чей жизненный цикл полностью или почти полностью может осуществляться в любой из двух зон (например, Масота balthica, Arenicola тarina — литоральные и сублиторальHbe, o HroxeTL Pelos colex benedeni, Clitellio arenarium, KJ1 e Halocarus hexacanthиs — литоральные и супралиторальные); б) виды, одна или несколько стадий жизненного цикла которых обязательно проходят на литорали, а остальные — за ее пределами (Han pH Mep, Nereis virens, Asterias rubens).
3. Виды, периодически посещающие литораль (например, Виссіпит ипdatит, голожаберные моллюски Cadlina laeuis, Coryphella liпеata, Eиbгапс?иs exigииs и др.); неизвестно, в какой степени этот биотоп обязателен для протекания их жизненного
HK|
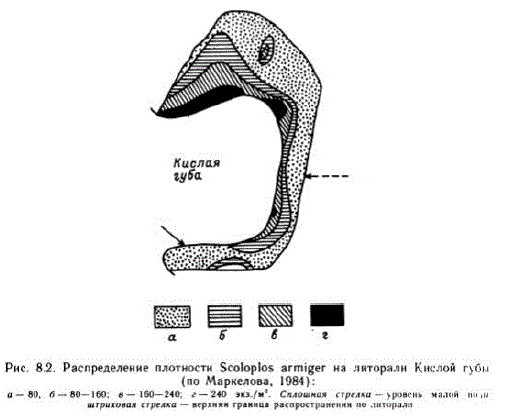
Сложность абиотической ситуации на литоралии особенности экологии видов сказываются на распределении биоты, поражающем своей мозаичностью. Но, как и в других приливных морях, в Белом море давно отмечена регулярность этого распределения — его поясность, обусловленная существованием обсуждавшегося выше широтного вектора (Гурьянова, Ушаков, 1929; Гурвич, 1934; Абрикосов, Соколова, 1948; Соколова, 1957; Луканин, Ошурков, 1981 и др.). Поясность наиболее ярко выражена у основных форм флоры (фукоиды). Регулярность распределения фауны также хорошо известна, однако она часто нарушается. Наиболее наглядно это проявляется в распределении макрофауны. Пространственная структура популяций, как правило, складывается из фона, часто весьма рассеянного, и агрегаций — ядер. Ядра могут представлять собою сплошную и длинную ленту, приуроченную к определенному горизонту литорали, как это часто наблюдается, например, у Ѕетibalапиs balanoides," или лентовидные отрезки, резко смещенные один относительно другого (Jaera). Чаще же ядра — это отдельные скопления, ориентированные главным образом вдоль линии урезаводы, которые могут образовывать скопления второго порядка с той же ориентацией — например, y Scoloplos armiger, Macoma balthica, Hydrobia uluae (Cakp, 1981; Маркелова, 1984) (рис. 8.2). У малоподвижных и неподвижных форм взаимное положение ядер и фона сравнительно стабильно в пространстве и времени, у подвижных — динамично. Положение ядра и особенности фона в значительной мере зависят от характера осадка, однако никогда не определяются только им, но комплексом факторов, в котором преимущество получают то один, то другие.
Известно, что макрофауна разделяется на обитающую на грунте и над грунтом (онфауна) и обитающую в грунте (инфауна) (Зенкевич, 1951, 1970). Приставка «эпи-» (например, эпифауна, эпибиоз) все чаще употребляется там, где речь идет о фауне обрастаний — малоподвижной и сессильной. В связи с этим нам представляется целесообразным ограничить теряющий определенность термин «онфауна», обозначив им только вагильный бентос. аким образом, по отношению к субстрату мы выделяем три группы — эпи-, ин-ионфауну. В расселении эпи- и инфауны большую роль играет личиночная стадия. Оседание личинок — равномерное или агрегированное — происходит при наличии подходящего субстрата и времени, достаточного для осуществления ком|- лекса поведенческих реакций, иногда на таких участках, где дальнейшее развитие проблематично или невозможно: например, оседание в мертвой зоне, мидий — на нитчатки заиленной литорали, балянусов — на валуны, обдираемые льдом, личинок седентарных полихет — в ванночкина скалнстой литорали и т. п. 3.1H Miiнация на ранних стадиях развития и роста постепенно создаг окончательную структуру поселения, как пространственную, так и возрастную. Так, В. В. Луканиным и В. В. Ошурковым (1981) выделены три типа поселений мидий на литорали, каждый ни которых характеризует определенный тип возрастных пирамн.1- Первый из них, при котором до 96 % составляют неполовозрелые особи, свидетельствует об экпоненциальном росте. Второй характ" ризуется равномерным распределением возрастной структуры. При третьем типе элиминация преобладает над пополнением, доминирующей группой оказываются половозрелые моллюски. При фор" мировании окончательной структуры потери могут быть очень велики. Так, по данным Н. М. Таракановой, на песчано-валунной литорали доля перезимовавших балянусов составила в 1978 Г. 4.8%, а в 1979 г. 7.5 %.
Эпифауна — в основном фауна твердых субстратов — заселяет в первую очередь нижний и средний горизонты литорали, а на прибойных участках поднимается и вверхний горизонт. Она может быть приподнята над дном настолько, насколько позволяют слой воды, покрывающий литораль, и элиминирующее воздействие ледового покрова. Оно и прибой, препятствующий осаждению личинок, рассеивающий планктон и вымывающий детрит со скалистойл иторали — пo- видимому, OHOBH факторы, ограничивающие развитие эпифауны там, где эта группировка преобладает. Поэтому наибольшая плотность и биомасса отмечены на более защищенных типах литорали — каменистой, валуннопесчаной. По мере заиления дна эпифауна лишается необходимого для нее субстрата — на валунно-песчано-илистой литорали она немногочисленна, а при отсутствии твердых фракций вообще HC LIP3 a EPT,
Инфауна - обитатели толщи грунта, глубина их распространения — до 40 см в зависимости от характера грунта. Виды инфауны заселяют всю литораль, избегая верхнего горизонта. Верхняя граница их расселения редко идет сплошным фронтом, чаще это языки, так как граница следует за увлажненными участками. Виды, доминирующие на песчано-илистой литорали, Arenicola тariпа и Масота balthica, обнаруживают распределение, близкоек равномерному. На промежуточных типах литорали усиливается агрегированность поселений. Так, например, поселения пескожила в этих случаях имеют лентовидный или спорадический характер, повторяющий распределение песка — предпочтительной фракции осадка (Калякина, 1971).
Онфауна — подвижная, способная в большей или меньшей степени избегать неблагоприятных условий группа видов, личиночная стадия не является необходимостью (правило, имеющее исключения). Онфауна расселяется повсем горизонтам литорали. Молодь на ранних стадиях ростаможет образовывать самостоятельные поселения, например литторины на нитчатках, но затем перемещается в биотопы, свойственные взрослым. Животные либо ведут убежищный образ жизни, либо защищены раковиной, которая не уменьшает их подвижности. Могут подниматься по камням вверх и неглубоко зарываться в грунт.
Этот краткий обзор экологических особенностей представителей макрофауны свидетельствует о возможности комбинаций ядер разных видов на разном фоне хотя бы только потому, Что виды занимают разные пространственные ниши (разобщены По толщине литорали). Распределение макрофауны в приливотливной зоне в первую очередь концентрирует внимание исслеКователей на роли абиотических факторов.
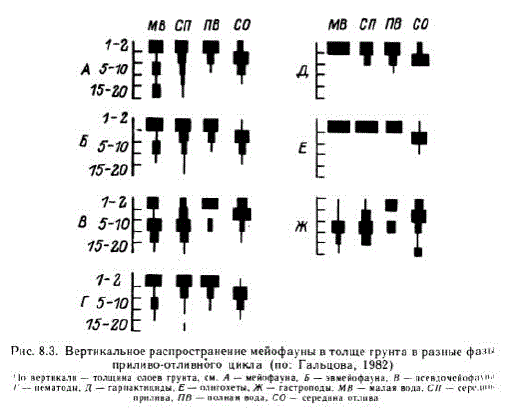
Размеры представителей мейо- и микрофауны определяют иной, нежели у макрофауны, масштаб распределения; например, вертикальная протяженность сообществ может измеряться уже не мет: и десятками сантиметров, а миллиметрами. Совместное сущевование нескольких видов в этих условиях неизбежно сопровождается их тонкой послойной стратификацией. Она описана В. В. Гальцовой (1982) у мейофауны. Видно, что разные группы входящие в состав мейобентоса, имеют свои характерные особен. ности распределения (рис. 8.3). Часть групп — остракоды, тур. беллярии, тихоходки, полихеты и молодь двустворчатых моллюс, ков — не обнаруживает вертикальных перемещений, чем отлича. ется от нематод, гарпактицид, олигохет и молоди брюхоногих моллюсков. Распределение четырех массовых видов гарпакти. цид в занимаемом ими верхнем 4-миллиметровом слое осадка связано с особенностями поведения рачков. Нипteтаппia jadensis распространена по всему слою практически равномерно, а Heterolaophonte minuta, Tachidius discipes H. Sten chelia palustris предпочитают верхний слой грунта. При этом Т. discipes во время прилива свободноплавает, а в отливоседает на влажный грунт, H. тіпи(а заякоривается на дне каудальными ветвями и щетин. ками, а S раIиstris активно ползает по поверхности грунта“. Такая комбинация распределения и поведенческих реакций позволяет этим животным избежать трофической конкуренции и достичь возможно более полного использования пищевых ресурсов ограниченного пространства, в котором к тому же повышается роль продуктов метаболизма как фактора среды.
Сообщества макрофауны, которая, по мнению ряда исследователей, вообще не дает примеров существования отдельных видовых популяций, в значительной мере регулируются эндогенно, Как показал И. В. Бурковский (1984), можно выделить некоторое минимальное пространство, которое характеризуется всеми свойствами данного биотопа и всем набором видов данного сообщества. Этот набор складывается из немногих доминирующих видов, на долю которых приходится преобладающая часть биомассы, и большего или меньшего количества второстепенных и случайных. Он сохраняется по мере увеличения пространства; сохраняется и доминирующая группа видов. Неоднородность увеличивающегося пространства обнаруживается в изменении порядка доминирования видов переходе сообщества от одного локального состояния к другому (рис. 8.4), сопровождающемся изменением количественных показателей (табл. 8.3). Резкое изменение среды вызывает полную смену доминантов.

Рассматривая литоральное население в целом, нельзя не отме. тить две сопряженные тенденции: континуальность, одна из при. чин которой заключается в широком, почти повсеместном распространении массовых форм, преодолевающем даже островную изо. лированность (транспортирующим средством в этом случае служат плавающие водоросли), и дискретность, которая выража. ется в реальности локальных биоценозов,
Субъединицы литорального континуума выделяют чаще всего по хорологическому признаку — таковы сообщества литорали разных морфологических типов, литоральных ванн, зоны выбросов, эстуариев. К настоящему времени накоплены и продолжают накан, ливаться характеризующие их количественные данные (Федоров, 1928; Гурьянова, Ушаков, 1929; Гурвич, 1934 Гурвич, Матвеева, 1939; Абрикосов, Соколова, 1948: Воскресенский, 1948: Броцкая, 1954; Соколова, 1957; Перцов, Флинт, 1963, и др.).
Не менее реальны и функциональные комплексы. Примером их может служить комплекс деструкторов растительного веще. ства. Он соответствует данному Ю. Одумом (1975) описанию ключевой группы деструкторов растительного опада и включает около десятка видов онфауны родов Littorina, Hydrobia, Gаттаrus, Marinogam marus, Jaera. Ha y HacTKax, oTJ1 Hua KOLLIHX CH C.To:}+K ным рельефом, рассеивается часть отмерших макрофитов. Детрит водорослевого происхождения проходит здесь все этапы трансформации от только что оторванного слоевища или его части до формирования зрелой (кондиционированной) разновидности. Деятель ность ключевой группы начинается на еще живых или только что оторванных растениях, наиболее старые части которых всегда повреждены и некротизированы. Мертвую ткань с ассоциированной микрофлорой потребляют литторины. Варьируя скребущие движения радулы, они очищают поверхность и выскабливают глубжележащие ткани гниющих водорослей или их крупных фрагментов (мелкие для питания неудобны, ибо они сдвигаются и свертываются под тяжестью тела моллюсков). Менее глубоко некротизированные макрофиты потребляются изоподами с их слабым ротовым аппаратом. Иногда в основании кустиков можно обнаружить целые колонии этих мелких рачков. Бокоплавы либо удерживают куски растений гнатоподами, либо цепляются за них, обкусывая края. Для их питания неудобны нефрагментированные слоевища, в особенности со слабо поврежденными покровами, Все названные животные, а также Hydrobiа иluае, могут питаться и кондиционированным детритом, оседающим на поверхности литорали. Мелкие моллюски и Jaera часто концентрируются на различных детритонакопителях.
Селективность выбора пищи обнаруживается только в последовательном включении видов ключевой группы в процесс трансформации макрофитов. В отсутствие детрита предпочитаемого размера все они могут питаться коагулирующим или сорбирующимся растворенным органическим веществом с ассоциированной микрофлорой, просто микрофлорой, фекалиями других видов и собственными, обрывками животных тканей и т. п. Большую оль в их питании играет и терригенная органика — вероятно, именно с нею связана высокая численность представителей этого комплекса в эстуариях.
Представители ключевой группы отличаются большей или меньшей подвижностью, что позволяет им быстро заселять биотопы с благоприятным сочетанием биотических и абиотических факторов. Условия среды могут изменяться, в частности, из-за приливо-отливной динамики в зависимости от направления ветра, сгонно-нагонных течений и других факторов очищаются от ила или заносятся им убежища бокоплавов и изопод, меняется расположение знаков ряби и, что важнее всего, рассеиваются по литорали частицы детрита. Бокоплавы активизируются в столбе воды высотою около 50 см; хаотическое движение рачков прекращается с повышением этого уровня, остальное время осушения или затопления они проводят в убежищах. Литторины и гидробии могут ползать как во время прилива, так восушной период пока на поверхности сохраняется влажная пленка. Пути изопод много короче, но для такого мелкого животного (максимальный размер 5.2 мм) на литорали существует много убежищ.
Контроль над обширной территорией приливо-отливной зоны осуществляется этими животными и чистомеханическим способом. Так, бокоплавы выносятся в зону выбросов водами сизигийных приливов и покидают ее с их прекращением. Литторины в массе забрасываются в самый верхний горизонтлиторали во время штормов, а затем самостоятельно спускаются вниз, проходя через богатую пищей зону выбросов. Гидробии переносятся водой, прикрепляясь к поверхностной пленке, изоподы — на кустиках водорослей, палках, иногда даже на мелких камнях. Естественно, эти перемещения сопровождаются частичной гибелью животных и некоторым ограничением сексуальных контактов, но именно таким образом осуществляется широкое расселение видов по литорали (фоновое) и формирование новых ядер.
Деструкторы образуют биоценозы и в более четких топических границах — там, где накапливается большое количество разнообразного детрита и гидродинамические условия таковы, что препятствуют сероводородному заражению. Все упомянутые виды в этом случае многочисленны: так, плотность Littorina Saxatilis может составлять до 400000 экз./м", в то время как на фоне она измеряется не более чем тысячами особей на 1 м". Здесь же наблюдается повышенная плотность паразитов и комменсалов. Такие биоценозы обязательно дополняют хищники — актинии, немертины, букцинумы, KJIeLL H Ta KHKE LOCTH Ta KOLLIL He высокой численности благодаря обилию пищи.
Функциональной характеристикой деструкторов является осуществляемый ими детритный круговорот. Он заключается в прохождении кооперированного органического вещества через единственное трофическое звенос выходом во внешнюю среду детритных новообразований — фекальных пеллетов. В ходе этого процесса могут иметь место следующие явления: 1) минерализация биогенов, в особенности дефицитного в высокоширотных морях азота; 2) гумификация органического вещества, которой способ. ствуют фенольные метаболиты фукоидов; 3) обогащение гумифицирующегося вещества непосредственно в кишечнике за счет бактериального синтеза; 4) обогащение его же за счет сорбции растворенного органического вещества и бактериального синтеза во внешней среде; 5) формирование устойчивых даже в гидродинамически активных условиях частиц детрита — фекальных пеллетов, которые в массе выносятся в сублитораль и глубже, 6) использование оформленных фекальных пеллетов или их фрагментов в качестве пищи сессильными или малоподвижными обитателями литорали или глубжележащих зон и их очередное преобразование вплоть дополной минерализации лабильных и захоронения в осадок стабильных компонентов органического веще(тв н.
Функция поддержания круговорота — не специфическая особенность ключевой группы. Ее отличает только полнота вовлеченного органического вещества, разнообразие способов питания, широкое распространение входящих в нее видов и динамичность пространственной структуры.
Остальная макрофауна литорали, за исключением хищников, питается более мелкими частицами детрита. Лишь небольшое число видов можно отнести к определенной трофической группировке: сестонофаги — мидии и балянусы, безвыборочные грунтоеды — пескожил и Capitella capitata. Большинство представителей макрофауны — полифаги. Так, Масота bal?tica и Fabricia sabella питаются и как сестонофаги, и как детритофаги, Phyllodoce тасиІata, EиІalia viridis — как плотоядные и детритофаги. Вся нехищная макрофауна тем или иным способом кооперирует органическое вещество, из которого потребляется только часть, другая же в виде фекалий, псевдофекалий, агглютинатов вновь оказывается во внешней среде.
Очевидно, что детритные круговороты осуществляются и другими представителями фауны приливо-отливной зоны. Вместе с тем на литорали существуют и гораздо более сложные функциональные комплексы, нежели рассмотренные выше. Для них характернатонкая селекция пищи, наличие нескольких трофических уровней, закономерные сукцессии. Примером такого комплекса может служить сообщество простейших (Бурковский, 1984). Основная пища инфузорий на морском дне — бактерии и микроводоросли. Пищевая специализация обнаруживается не только в предпочтении определенных видов пищи, но и в выборе размеров пищевых частиц. В морском псаммофильном сообществе, включающем 12 видов, где пищей служат в основном водоросли, две пары видов продемонстрировали предельно высокую степень перекрывания пищевых спектров (0,94—0,99), что свидетельствует об избытке соответствующего корма. Низкие коэффициенты перекрывания, свойственные остальным видам, обнаруживают лимит пищи и расхождение видов во времени и пространстве. Виды доминирующего комплекса комплементарны — три из них питаются водорослями разных видов, а четвертый — бактериями. В условиях постоянного притока пищи, который наблюдается в течение лета, трофическая структура биоценозаостается почти неизменной, тогда как видовой состав и, в особенности, состав доминирующего комплекса сильно варьируют.
Часть продукции инфузорий используется животными высших трофических уровней, но в сообществе достаточно велико содержание хищников, питающихся более мелкими инфузориями, что способствует возвращению в него существенной (до 40 %) части продукции. Одновременно сообщество простейших со всей его сложной структурой участвует и в общем детритном круговороте.
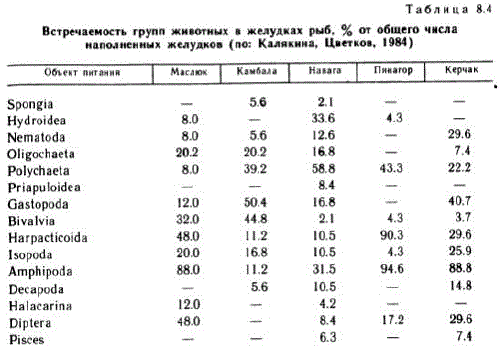
В целом население литорали в общей биологической системе моря является блоком, мобилизующим, перерабатывающим и подготавливающим к транспортировке в глубжележащие районы разнообразное по своей структуре автохтонное и аллохтонное (терригенное) органическое вещество. Отдельные представители литоральной фауны и флоры являются звеньями трофических цепей, уходящих за пределы осушной полосы. В приливо-отливной зоне кормятся птицы — как морские, так и не связанные непосредственно с литоралью и морем (Виноградов, 1950; Перцов, 1963, Перцов, Флинт, 1963, Бианки и др., 1979. Полищук и др., 1979, Бойко, Бианки, 1981, и др.). Мелководья Белого моря служат местом нагула для многих бентосоядных рыб (Азаров, 1963, Калякина, Цветков, 1984). Н. М. Калякиной и В. И. Цветковым непосредственно на литорали обнаружено 18 видов рыб, из которых 8 приходят на литораль в конкретные сроки или постоянно отлавливаются в определенных стациях: это полярная камбала, керчак, ледовитоморская рогатка, девятииглая колюшка, маслюк, навага, пинагор, беломорская треска (Liopsetta glacialis, MyохоCephalus scorpius, M. quadricornis, Pungitius pungitius, Pholis gunnelis, Eleginus navaga, Cyclopterus lumpus, Gadus morhua maris-albi.).
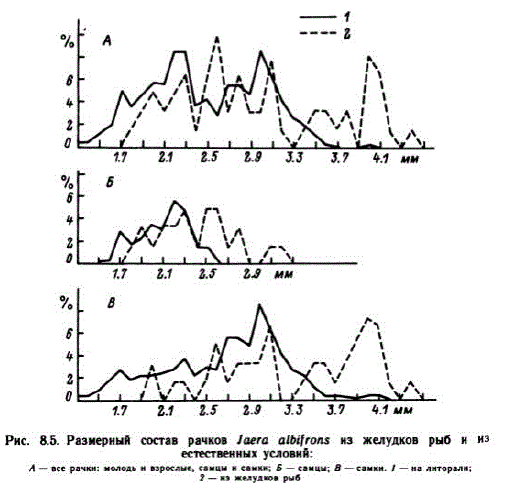
Наблюдения показали, что рыбы, приходящие в светлое время суток на литораль, по-разному осваивают ее. Камбала идет вместе с приливной водой в слое воды 40 см и более, держится небольшими группами по 5—12 экземпляров, особи обычно близки по размерам. Рыбы проплывают 2—5 м и затаиваются на дне, либо движутся над ним, приподнимая головой рыхлый осадок. Годовики появляются на литорали в августе, держатся группами, часто вместе с сеголетками. Двухлетние особи приходят только в темное время суток. В конце августа в группах наваг преобладают двухлетки, многочисленны и годовики, сеголетки единичны. С появлением закраек льда двухлетние наваги, а часто и треска, идут кор-" миться сюда вместе с приливной водой, надламывающей лед. В сентябре на первом месте по численности оказались сеголетки пинагоров и керчаков, среди маслюков годовики и сеголетки были представлены поровну. Пинагоры держатся зарослей фукоидов и в прилив наведываются даже в редкие куртины верхней трети литорали. Вероятно, особенностями поведения рыб можно объяснить селективный отбор некоторых видов пищи. Так, из сравнения возрастно-полового состава Jaera albifrons, обитающих на литоралии извлеченных из желудков рыб, обнаружилось, что рыбы выедают наиболее крупных самок и самцов (рис. 8.5). В табл. 8.4 приведены сведения о частоте встречаемости разных объектов питания в желудках рыб.
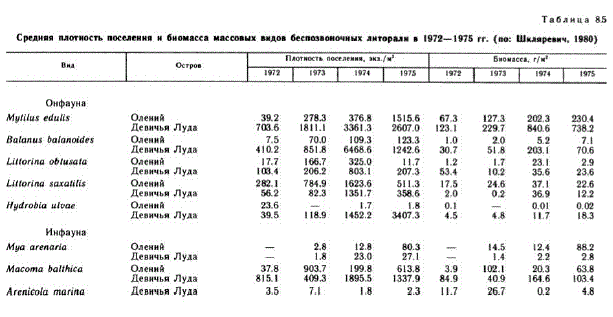
Из известных объектов марикультуры (Муtilus edиlis, Mya arenaria, Obelia longissiта) в Белом море культивируются мидии (Кулаковский, Кунин, 1983). Представители литоральной фауны не являются обязательным компонентом пищи рыб. Однако, как было показано выше, рыбы систематически подходят к берегам и некоторая их часть заходит для откорма в приливо-отливную зону. Таким образом, население литорали может рассматриваться как запас пищи, а также и как объект марикультуры при искусственном рыборазведении (наиболее вероятны капшаки-бокоплавы). В этой связи сравнительно малоизученная проблема — динамика численности массовых видов беспозвоночных литорали — обретает большой практический и научный интерес. В работе Г. А. Шкляревич (1980) приведены данные о нескольких массовых видах, плотность поселения которых изучалась в 1972— 1975 гг. (табл. 8.5). Причины, по которым некоторые из них обнаружили тенденцию к увеличению обилия, разнообразны: для Hydrobiа иluае это увеличение на литорали нитчаток в результате штормового нагона, для Mya arenaria — восстановление зостеры, изменяющей структуру грунта, и т. п. Увеличение численности и биомассы других видов Г. А. Шкляревич объясняет высокой среднелетней температурой 1972—1974 гг. Можно думать, что среди факторов, влияющих на численность видов, для которых Белое море по гидрологическим условиям является северным краем ареала, температура занимает одно из первых мест. Способность использовать местные и временные потепления для обеспеLIEHFH размножения H ранних периодов роста H3BeTH Heкоторых видов (Бек и др., 1980). Механизмы, обусловливающие реакцию видовой популяции на температуру, разнообразны — это или удлинение сроков размножения, прямо обеспечивающее более высокую численность потомства (Масота balthica), или, напротив, их сокращение, благодаря чему часть теплого сезона высвобождается для роста сеголеток, что в свою очередь способствует снижению смертности на ранних стадиях (Jaera). Отмечен активный поиск теплых линз в поверхностном слое воды личинками Asterias rubens и сосредоточение личинок пескожила в верхнем наиболее прогреваемом горизонте литорали. У бокоплавов результатом теплоголета является увеличение доли самок в потомстве, что позволяет ожидать увеличения численности потомства на следующий год.
Вместе с тем следует отметить, что благоприятное для многих видов макрофауны повышение температуры иногда приводит к серьезным нарушениям в более сбалансированных сообществах мейо- и микрофауны, что может вызвать общее ухудшение условий обитания и сказаться на характере протекающих в этой зоне процессов деструкции.
Следует отметить, что Белое море — водоем, в котором, по подсчетам Е. Н. Черновской (1956), на 1 км" зеркала приходится около 30 м берега (без учеталуд, корги островов). Роль прибрежной зоны, таким образом, оказывается более значительной, чем в любом другом водоеме России. В ней сосредоточено около 1.5 млн т живой массы макрофитов, от которой ежегодно отторгается и подвергается переработке не менее 1 млн т. Прижизненные и посмертные выделения растений и, по-видимому, животных (см., например: Кулаковский, Кунин, 1983) резко повышают содержание в прибрежной зоне растворенного органического вещества, а его поверхностно-активные компоненты, сорбирующиеся на органическом субстрате, способствуют развитию микрофлоры. Коагулирующее или сорбирующееся на минеральных частицах высокомолекулярное растворенное органическое вещество, как это показали для баренцовоморского побережья В. Е. Стрельцов и И. Я. Агарова (1978), может в течение одного приливо-отливного цикла трансформироваться в минеральные и низкомолекулярные формы биогенов, причем среднегодовое их количество, по оценке авторов, составляет около 1000 и 40 кгс 0.15 км" литорали с поселением фабриции и пескожила. Литоральная зона — это зона бурного развития эпифитов (Бондарчук, 1974). Зоо- и фитопланктон также обнаруживают более высокую плотность в прибрежной зоне. Это разнообразное органическое вещество пополняется и за счет гибели бентосных животных — окраска бенгальским розовым TOHKOTO литорального LLLLLL LLLLLLLL0LLLL LLLaL LLLLLaLLL присутствие в нем деструктурированного свежего органического вещества животного происхождения. Наконец, через литоральную зону стекает терригенное органическое вещество. Здесь оно задерживается штормовыми выбросами, валунно-каменистым барьером нижней литорали, оседает в литоральных ваннах и депрессиях поверхности и затем смешивается с автохтонным органическим BELIL EPCTB (O M.
Мобилизованное и переработанное сообществом литорали до мелкодисперсной-массы или оформленное впеллеты и агглютинаты органическое венцество уносится в глубжележащие зоны (Лисицын, 1983), где продолжается детритный круговорот, поддержи. вающий население сублиторалии батиали, возможно, в не мень, шей степени, чем это осуществляется через пастбищные пищевые |I?:TIH.
Роль детрита как источника пищи, постоянным и непрерыв. ным потоком поступающего в экосистемы и тем самым обеспечивающего их буферность, хорошо известна (Остапеня, 1979). Однако количественные характеристики этого потока изучены значительно хуже, нежели те, что позволяют оценить приток органи. ческого вещества через пастбищные цепи. В первую очередь это связано с методическими трудностями, преодоление которых требует усилий специалистов разных профилей. Оценка потока органического вещества в детритных круговоротах и детритных пищевых цепях — одна из основных задач современных исследо. ваний в прибрежной зоне моря.



