Макробентос - Фитобентос
Наличие в Белом море большого числа мелководных, хорошо прогреваемых и освещенных участков создает благоприятные условия для обитания различных макрофитов. Крайне важно также, чтобы на дне преобладали каменистые субстраты, к которым водоросли крепятся своими ризоидами.
Больше всего таких участков в прибрежных районах, в многочисленных островных архипелагах Кандалакшского и Онежского заливов. В остальных частях акватории водоросли встречаются реже и в меньшем количестве.
Всего в Белом море насчитывается 185 видов донных макрофитов. Больше всего среди них бурых водорослей – 75 видов. Среднюю позицию занимают зеленые водоросли – 69 видов. Беднее других флора беломорских багрянок – 39 видов. К числу макрофитов Белого моря принадлежат и 2 вида высших водных растений: Zostera marina и Eleocharis sp. (Возжинская, 1980; Возжинская и др., 1995).
Вертикальное распределение макрофитов в Белом море закономерно изменяется по мере продвижения от супралиторали в глубину.
Во многих местах по берегам моря часто встречаются скопления гниющих макрофитов*, выброшенных волнами. Их основу составляют фукоиды, часты выбросы анфельции. Слоевища ламинарий и (или) их куски встречаются в выбросах значительно реже. Они обычно опускаются на дно и сносятся на глубины. Валы штормовых выбросов макрофитов в ряде случаев достигают 10 м в ширину при высоте до 2 м. Их масса варьирует в широких пределах: от 1.5 до 5 кг/м2 (Бек, Глазов, 1995; Гришанков, Нинбург, 1995). Источником формирования этих скоплений служат плавающие водоросли, сначала
оторванные от субстрата и проплававшие некоторое время, а затем выброшенные волнами на берег. Основу их биомассы составляют Fucus vesiculosus, F. serratus, Ascopyllum nodosum, Chorda fillum и Chordaria flagelliformis (Халаман, Бергер, 2006). В Кандалакшском заливе 85% от биомассы плавающих объектов дают фукоиды. В Онежском заливе на их долю приходится 41%, а на долю хорды и хордарии – 51% от общей биомассы. На основании полученных данных (табл. 27) подсчитано, что единовременно в Белом море обнаруживается около 42 тыс. т плавающих водорослей и сопутствующих им животных (мидий, литорин, полихет и др.). При этом на долю перечисленных выше бурых водорослей приходится от 85 до 92% от общей биомассы плавающих объектов, т.е. приблизительно 37 тыс. т влажной массы.
Таблица 27 Биомасса и плотность распределения плавающих объектов в разных частях Белого моря (по: Халаман, Бергер, 2006)
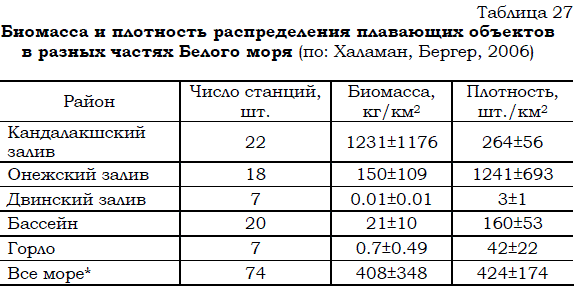
* Без Воронки и Мезенского залива.
Большая часть макрофитов произрастает на литорали и в верхней сублиторали. Для мест с вертикальной стратификацией вод характерно поясное распределение макрофитов, сменяющих друг друга по мере продвижения в глубину. При этом, наряду со сменой видового состава и доминирующих видов, изменяются и количественные показатели их обилия. Основу биомассы макрофитов в поселениях, располагающихся на каменистых грунтах от литорали до 1.5–2 м ниже нуля глубин, составляют различные фукоиды: Fucus vesiculosus, F. serratus, Ascopyllum nodosum и др. При 100%-ном покрытии дна их биомасса достигает 4–5 кг/м2, но в большинстве случаев не превышает 1–2 кг/м2.
На песчаных и илисто-песчаных грунтах вдоль Карельского и Поморского берегов на этих же глубинах обнаруживаются заросли морской травы Zostera marina L. В 1960 г. беломорская зостера вымерла по всему морю, подобно тому, как это произошло в 30–40-х годах прошлого столетия в морях Атлантики, но тогда не затронуло Белое море. Наиболее вероятной версией гибели этого растения считается поражение паразитическим миксомицетом Labirintula macrocystis Cienk. (Fischer-Piette et al., 1932; Hunstmann, 1933; Blegvad, 1934; Butcher, 1934; Petersen, 1934; Renn, 1936; МорозоваВодяницкая, 1939; Dexter, 1950 и др.).
В периоды наибольшего обилия биомасса зостеры достигала 6 кг/м2 (Гемп, Паленичко, 1956; Гемп, 1962,1969). В наши дни ее заросли медленно восстанавливаются, хотя биомасса зостеры пока не достигла прежних величин и не превышает 1–2 кг/м2 (Вехов, 1995).
На каменистых грунтах глубже пояса фукоидов (от 2 до 8 м) располагаются заросли макрофитов, в которых преобладает Laminaria saccharina. При сплошном покрытии дна средняя биомасса водорослей в этих донных биоценозах может достигать почти 4 кг/м2, однако обычно поселения макрофитов на этой глубине в той или иной степени разрежены, а показатели их обилия бывают гораздо более низкими. Далее по мере продвижения в глубину L. saccharina постепенно исчезает. Сначала ее замещает близкий вид L. digitatа, а затем – красные водоросли. Биомасса макрофитов на этих глубинах не превышает 10–30 г/м2. Глубже 22 м макрофиты в Белом море не обнаружены (Денисов, Денисова, 1979).
В тех местах, где из-за повышенной гидродинамики «бассейновая структура» вод нарушена, описанное выше поясное распределение макрофитов отсутствует. Подтверждением этому может служить тот факт, что в Онежском заливе заросли ламинарии встречаются на разных глубинах, начиная с минимальных: сразу от перехода литорали в сублитораль (Возжинская и др., 1995).
Запасы макрофитов в Белом море начали исследоваться около 80 лет назад в основном в связи с необходимостью удовлетворить сырьевые потребности производства сначала йода, а позже и других продуктов переработки водорослей: агар-агара, альгинатов и маннита. Первые данные были сильно завышены. Предполагалось, что одни только штормовые выбросы могут давать ежегодно порядка 1650 тыс. т макрофитов (Марциновский, 1930).
Первые достаточно надежные материалы наблюдений свидетельствовали о том, что запасы водорослей составляют приблизительно 1.5 млн. т сырца (Милоградов, 1938; Чернов, 1945). В дальнейшем водолазными работами, ландшафтным картированием и другими методами запасы водорослей были уточнены. По состоянию на 1984 г. биомасса промысловых макрофитов (без зостеры) составляла около 1 млн. т (табл. 28).
Таблица 28 Запасы промысловых водорослей (тыс. т влажной массы) в Белом море (с изменениями по: Сорокин, Ванюхина, 1995)
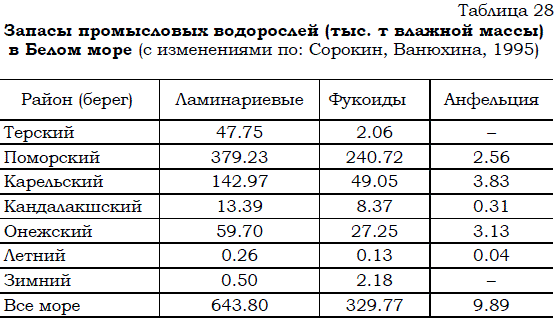
По сведениям из других источников (Макаров, 1998) спустя примерно 20 лет запасы макрофитов составляли сходную величину порядка 1 млн. т, хотя доля ламинарий была больше, чем указывалось ранее, а обилие фукоидов, наоборот снизилось (табл. 29).
В середине 90-х годов прошлого века сотрудники СевПИНРО (Зеленков, Стасенков, 1997; Пронина, 1998) оценили запас ламинариевых водорослей величиной в 500– 550 тыс. т влажной массы, фукоидов – в 250–300 тыс. т, а
анфельции – в 6 тыс. т. В 2003 г. результаты наблюдений сотрудников этого же учреждения показали, что общий запас ламинариевых водорослей составлял 600–750 тыс. т сырца, а запас фукоидов был равен 300–350 тыс. т (Пронина, 2001; Отчет СевПИНРО, 2003).
Таким образом, можно считать установленным, что запас макрофитов в Белом море равен приблизительно 1.5 млн. т влажной массы, включая и зостеру, запас которой составляет около 400 тысяч тонн.
Таблица 29 Запасы промысловых водорослей (тыс. т) в обследованных районах Белого моря (с сокращениями по: Макаров, 1998)
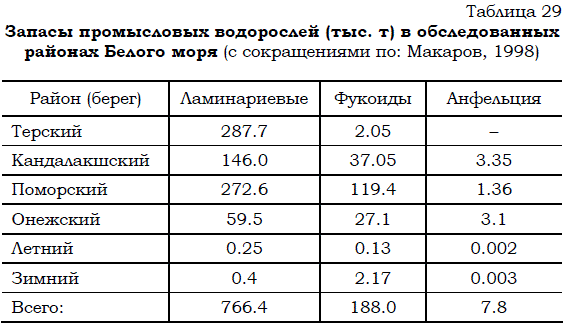
Рост и развитие макрофитов в Белом море начинаются в марте еще подо льдом. При этом уровень фотосинтеза остается крайне низким (Возжинская, 1971). В дальнейшем после освобождения моря ото льда, по мере увеличения уровня солнечной радиации скорости роста и фотосинтеза увеличиваются в десятки и даже сотни раз. При этом происходит резкое увеличение биомассы макрофитов. Пик этих процессов приходится обычно на июнь. Летом (июль – август) при созревании водорослей процессы роста и фотосинтеза замедляются. Лишь осенью вновь наблюдается некоторая активация фотосинтетических процессов, уровень которых, однако, не достигает величины весеннего максимума. При этом начинается отмирание вершины таллома у ламинарий и генеративных ветвей фукоидов. Огромные массы органического вещества поступают в море в виде кусков макрофитов, включаясь в поток детрита. Биомасса водорослей начинает снижаться.
Экспериментальных исследований продуцирования у беломорских макрофитов практически нет. Была выполнена только одна работа, но и ее результаты представляются весьма сомнительными (Возжинская, 1971). Согласно этим данным весной в период наибольших роста и развития за сутки синтезируется: фукусами от 82 до 115 г С/м2 площади зарослей, аскофиллумом – от 134 до 145 г С/м2 и ламинарией сахаристой – от 90 до 142 г С/м2. За весь вегетационный период суточные показатели продукции у этих макрофитов в среднем составляют соответственно 22, 19 и 26 г С/м2. Если пересчитать на год, приняв длительность периода вегетации равной таковой для фитопланктона, т.е. 180 суткам, то получится, что в среднем макрофитами продуцируется около 4 кг углерода на 1м2 площади зарослей макрофитов. Позже В.Б. Возжинская (1986) получила более низкие средние величины годовой продукции беломорских макрофитов: 1.1 кг С/м2. Однако, несмотря на то, что эти последние данные были почти в 4 раза ниже первоначальных, они все равно представляются завышенными, поскольку слишком сильно отличаются от результатов иного расчета продукции, которые будут приведены ниже.
Исходя из данных В.Б. Возжинской (1986), можно рассчитать величину годовой продукции макрофитов следующим образом. При средней биомассе около 3 кг/м2 (Сорокин и др., 1984) и запасе макрофитов около 1.5 млн. т, занимаемая ими площадь составляет порядка 5.108 м2. Следовательно, при указанных выше величинах суточной продукции и длительности периода вегетации, годовая продукция составляет около 0.57.1012 г С, что энергетически эквивалентно 0.57.1013 ккал. От уровня продукции фитопланктона это составляет около 10%, что на порядок больше аналогичных показателей, характерных для продукции макрофитов в Мировом океане (Моисеев, 1989). Таким образом, данные о продукции макрофитов, использованные в приведенном выше расчете, по-видимому, сильно завышены.
Результаты расчета продукции макрофитов с помощью второго метода дали иные результаты. Принято считать, что годовой P/B-коэффициент макрофитов равен приблизительно единице (Алимов, 1989). Иными словами водоросли производят за год массу, равную исходной, т.е.
удваивают ее. Следовательно, годовая продукция макрофитов в Белом море составляет примерно 1.5 млн. т сырого вещества, калорийность которого равна 0.5 ккал/г (см. табл. 1). Значит, годовая продукция этих макрофитов в Белом море составляет порядка 0.75×1012 ккал. Эта величина почти на порядок меньше результатов расчета, произведенного на основании экспериментальных данных о суточной продукции макрофитов. По-видимому, до получения дополнительных материалов целесообразно в дальнейших расчетах пользоваться средней величиной, общей для всего моря годовой продукции макрофитов, равной 0.32.1013 ккал или около 35 ккал/м2.
В этом случае годовая продукция беломорских макрофитов составляет около 5% от продукции фитопланктона, если считать последнюю эквивалентной 6.0.1013 ккал, что также, хотя уже и не столь значительно, превосходит аналогичное соотношение в Мировом океане (Моисеев, 1989). Очевидно, что столь высокий уровень продукции макрофитов обусловлен наличием в Белом море больших площадей мелководных, хорошо прогреваемых и хорошо освещаемых участков, благоприятных для произрастания водорослей и их продуцирования.
При этом, однако, следует отметить, что продукция макрофитов, сколь бы значительной она ни была, играет совершенно иную роль, чем продукция фитопланктона. В Белом море практически нет организмов, питающихся макрофитами. Они могут входить в состав пищи в основном в виде кусочков полуразложившихся слоевищ, подвергающихся бактериальной деструкции. В таком виде они утилизируются морскими ежами и некоторыми моллюсками. По мере дальнейшего уменьшения размеров кусочков и увеличения доли их бактериальной переработки они становятся доступными для использования в пищу все большему количеству детритофагов, входящих в состав как пелагических, так и донных сообществ. В связи с этим очевидно, что достаточно высокая величина продукции макрофитов свидетельствует, в частности, о развитости в Белом море детритной цепи утилизации вещества и энергии.



